
Remember me
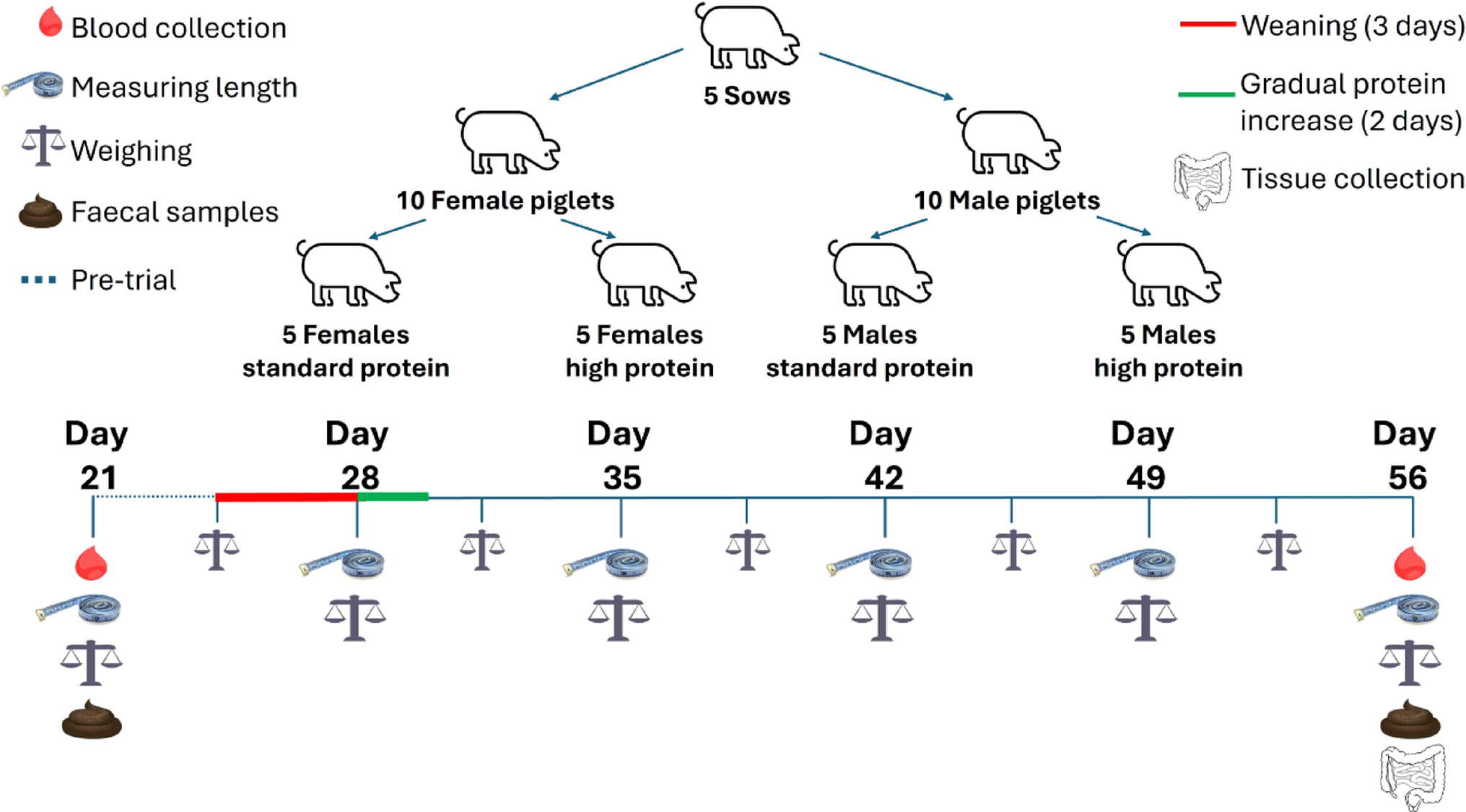
The twenty male and female piglets consuming a SP or HP diet were monitored weekly for changes in length and comparisons were made between protein group (Fig. 2a), sex (Fig. 2b), and protein group by sex (Fig. 2c). The changes in weight were measured bi-weekly and compared between SP and HP groups (Fig. 2d), males and females (Fig. 2e) and between the protein and sex interactions (Fig. 2f). Significant differences in length between the HP and SP groups started to appear at 28 days (p = 0.02), with weight differences emerging by 31 days of age (p < 0.001), persisting for the duration of the study (Fig. 2g). Differences in weight and length were observed exclusively under different protein conditions, with no significant differences between males and females or any interaction effects between sex and protein.
Fig. 2
Twenty piglets entered the centre for dairy research (CEDAR) at the University of Reading at 21 days old and fed sow replacer milk for three days before being gradually weaned onto standard weaner feed over the following three days. At 28 days old, the piglets were assigned into litter-matched and sex-balanced treatment groups and fed either a standard protein (SP; 18% dietary protein) or high protein (HP; 28% dietary protein) diet until 56 days old. Increases in body length were tracked to assess the effects of dietary protein a, sex b and the interaction between sex and protein: standard protein females (SP/F), high protein females (HP/F), standard protein males (SP/M) and high protein males (HP/M) c. Similarly, weight gain was measured to evaluate the influence of dietary protein d, sex e, and the interaction between sex and protein f. Significant differences between groups in changes from baseline were determined using the Kruskal-Wallis test with pairwise comparisons and Benjamini Hochberg corrections g. Results are presented as mean ± SEM and n = 5 piglets/sex/treatment group
Increased dietary protein intake influences alpha diversity but not beta diversityTo investigate the impact of dietary protein on microbiota diversity, alpha and beta diversity analyses were conducted on faecal samples from piglets subjected to high or SP diets over a four-week period. Faith’s PD remained stable in the HP group throughout the 5 weeks, while it significantly declined in the SP group (p = 0.037), resulting in lower diversity in the SP group than the HP group by the end of the trial (p = 0.047) (Fig. 3a). By day 56, the HP group also exhibited significantly more observed features compared to the SP groups (p = 0.049) (Fig. 3b). However, no significant differences between the protein groups were observed for Shannon diversity (p = 0.36) (Fig. 3c). There were no differences in any of the measured alpha diversity indices between males and females at any time point (Fig. 3d, e and f). A significant interaction effect between protein intake, sex, and day was detected for Faith’s PD (p = 0.028) (Fig. 3g), however, no significant differences emerged in pairwise comparisons after FDR correction. Additionally, no interactions were found between protein intake, sex, and day for observed features (Fig. 3h) or Shannon diversity (Fig. 3i).
Fig. 3
Twenty piglets were allocated into four litter-matched and sex-balanced treatment groups to consume either a standard protein (SP; 18% dietary protein) or high protein (HP; 28% dietary protein) diet for 28 days from 28 days old. Faecal samples were analysed by 16 S rRNA sequencing and the differences in alpha diversity between the HP and SP treatment groups at day 21 and 28 was assessed using Faith’s PD a, Observed features b, and Shannon diversity c. Similarly, differences in alpha diversity between males (M) and females (F) at days 21 and 56 irrespective of diet were measured by Faith’s PD d, Observed features e, and Shannon diversity f. The interaction between sex and diet on alpha diversity was also measured by Faith’s PD g, Observed features h, and Shannon diversity i. Statistical analyses were conducted using Kruskal-Wallis test with pairwise comparisons. Corrections for multiple testing were conducted using the Benjamini-Hochberg method
Weighted UniFrac analysis revealed distinct clustering patterns over the trial duration for each group but showed no variations attributable to dietary protein intake or sex (Fig. 4a and b). At the study’s onset (day 21), there were notable differences between HP/F and SP/F (p = 0.04), and between HP/F and HP/M (p = 0.02) (Fig. 4c). By day 56, these initial differences had dissipated (Fig. 4d). The Bray-Curtis dissimilarity analysis did not identify any significant differences between protein groups (Fig. 4e), males and females (Fig. 4f), or any sex-by-protein interactions at day 21 (Fig. 4g) or day 56 (Fig. 4h).
Fig. 4
The beta diversity of the faecal microbiota from female (F) (n = 10) and male (M) (n = 10) piglets consuming either a standard protein (SP; 18% dietary protein) or high protein (HP; 28% dietary protein) in a 2 × 2 model for 28 days. The data is presented as a weighted unifrac distance matrix comparing protein groups at days 21 and 56 a, sex at days 21 and 56 b, the interaction between protein and sex at day 21 c and day 56 d. The Bray-Curtis dissimilarity index was also used to compare SP and HP at days 21 and 56 e, males and females at days 21 and 56 f, the interaction between sex and diet at day 21 g and day 56 h. The p values were determined using PERMANOVA with pairwise comparison and Benjamini-Hochberg corrections. * = p < 0.05
Interaction between dietary protein intake and sex on microbiota composition and functionFaecal microbiota from piglets that consumed a SP diet or HP diet for four weeks were taxonomically profiled using 16 S rRNA sequencing. The differential abundance of the ASVs present were determined (Fig. 5a) and analysed using DESeq2. An analysis of how sex influenced the abundance of different microbial families and genera in piglets, regardless of diet, revealed that female piglets exhibited significantly elevated levels of several bacterial families, including Weeksellaceae (p < 0.001), Acrobacteraceae (p = 0.03), Xanthobacteraceae (p = 0.03), and Moraxellaceae (p = 0.03) (Fig. 5b). Additionally, the genera Chrysobacterium (p < 0.001) and Bradyrhizobium (p = 0.042) were also more abundant in females than in males (Fig. 5c). Further, when comparing sex within the dietary groups, a distinct interaction between sex and diet emerged. In the SP condition, there were 12 genera with significant differences between males and females, including the potentially pathogenic Eschericia-shigella (p < 0.001), which was more abundant in females, and the generally protective Bifidobacterium (p < 0.01), which was more abundant in males (Fig. 5d). Similarly, within the HP diet condition, multiple sex differences in bacterial abundance were observed, including a higher abundance of genera associated with disease states, Arthrobacter (p = 0.025) (Fig. 5e) Bradyrhizobium (p = 0.003) (Fig. 5f), Chryseobacterium (p < 0.001) (Fig. 5g), Paracoccus (p < 0.001) (Fig. 5h), Lawsonella (p = 0.011) (Fig. 5i), and Staphylococcus (p < 0.001) (Fig. 5j) in the faecal microbiota of females in comparison to males. Conversely, males exhibited elevated levels of Pyramidobacter (p = 0.007) (Fig. 5k), butyrate-producing Lachnospiraceae NK4A136 (p = 0.007) (Fig. 5l), and the potentially pathogenic Cloacibacillus (p = 0.016) (Fig. 5m). The presence of more sex differences in the abundance of bacterial genera within each of the diet groups in comparison to when the dietary groups are analysed together.
Fig. 5
Twenty piglets were litter-matched into either a standard protein (SP; 18% dietary protein) or a high protein (HP; 28% dietary protein) diet for 28 days in a 2 × 2 sex-balanced experimental protocol. The faecal microbiota of the piglets was analysed by 16 S rRNA sequencing to determine the relative abundance of bacterial families after 28 days on their respective diets a. Differential abundance analysis was performed using DESeq2 to determine log2 fold changes in bacteria families b and genera c between males and females. Additionally, differences in genera were analysed between standard protein-fed females (SP/F) and males (SP/M) d. Significant differences between high protein-fed males (HP/M) and females (HP/F) were determined using DESeq2 e. Multiple testing corrections were applied using the Benjamini-Hochberg method, with significance defined as an adjusted p-value < 0.05. Differential abundance analysis was performed using DESeq2 to determine log2 fold changes in bacteria families b and genera c between males and females. Additionally, differences in genera were analysed between standard protein-fed females (SP/F) and males (SP/M) d. Differences between high protein-fed males (HP/M) and females (HP/F) were determined using DESeq2 and those that reached significance are presented: Arthrobacter e, Bradyrhizobium f, Chryseobacterium g, Paracoccus h, Lawsonella i, Staphylococcus j, Pyramidobacter k, Lachnospiraceae_NK4A136 l and Cloacibacillus m. Multiple testing corrections were applied using the Benjamini-Hochberg method, with significance defined as an adjusted p-value < 0.05
Functional profiles of the microbiome were characterised using KEGG Orthologs (KOs), MetaCyc pathways, and the KEGG Orthology database. Comparative analysis between groups was conducted using DESeq2. Notably, females had a significantly elevated abundance of the MetaCyc pathways involved in carbohydrate fermentation processes such as Clostridium acidogenic fermentation and Butanoate fermentation, in comparison to males, whereas males had no upregulated MetaCyc pathways in comparison to females (Fig. 6a). Similarly, females had an increased abundance in KEGG pathways associated with colorectal cancer, lung cancer, and viral myocarditis (Fig. 6b) in comparison to males. Consumption of a high-protein diet resulted in a significantly lower abundance of 2-oxo-3-hexenedioate decarboxylase (p < 0.001) (Fig. 6c), an enzyme involved in the degradation of aromatic compounds, and a reduction in the MetaCyc pathway responsible for the cleavage of aromatic compounds (p < 0.001) (Fig. 6d) compared to the SP diet, irrespective of sex. Furthermore, females consuming a high-protein diet exhibited 497 KOs that were significantly upregulated compared to males on the same diet, while 99 KOs were more abundant in males than females within the high-protein group. Figure 6e highlights KEGG orthologs with the lowest adjusted p-values and a minimum log2 fold difference of 1.5 between the HP/F and HP/M groups.
Fig. 6
The functional abundance profiles of faecal microbiota in 20 piglets (n = 10 males, n = 10; females) after consuming either a standard protein (SP; 18% dietary protein) or high protein (HP; 28% dietary protein) diet for 28 days were determined using PICRUSt2. Differences between males and females were evaluated for MetaCyc pathways a and KEGG pathways b. The effects of dietary protein were assessed usng KEGG orthologs c and MetaCyc pathways d. Additionally, we analysed the interaction effects between dietary protein intake, and sex, also using KEGG orthologs e, KEGG pathways f and MetaCyc pathways g. Significances were determined using DESeq2 with Benjamini-Hochberg multiple testing corrections and was set at adjusted p-value < 0.05
High-protein diets enhanced the potential of the male microbiome to metabolise protein (p = 0.042) and produce lipopolysaccharide (p = 0.027) more than in females. Conversely, the female microbiome demonstrated upregulation of pathways linked to disease states such as bladder cancer (p = 0.043) and prion disease (p = 0.02) (Fig. 6f). Additionally, females on the high-protein diet showed an increase in pathways involved in caprolactam degradation (p = 0.026), a metabolite formed from the hydrogenation of phenol to cyclohexanone. The MetaCyc pathways also revealed an increased abundance of pathways associated with menaquinol (vitamin K2) production and biosynthesis of cob(II)yrinate a, c-diamide reductase, an enzyme involved in vitamin B12 synthesis, in females compared to males (Fig. 6g).
Dietary protein increased the production of p-cresol in a sex-dependent mannerTo measure the end-stage microbial metabolites produced from protein fermentation, urine samples were collected from the bladders of male and female piglets that had consumed either a SP or HP diet for four weeks and analysed using SPME-GCSMS and ammonia assay kits. A significantly higher urinary concentration of p-cresol in piglets fed the HP diets compared to those on the SP diets was observed (Fig. 7a) (p = 0.013), indicating that excessive dietary protein reached the colon for fermentation by the resident microbiota, which supports the hypothesis. However, no significant differences were noted in other metabolites including phenol (Fig. 7b), indole (Fig. 7c), or ammonia (Fig. 7d).
Fig. 7
Protein-associated urinary metabolites were excreted from male (M; n = 10) and female (F; n = 10) piglets consuming either standard protein (SP; 18% dietary protein) or high protein (HP; 28% dietary protein) diets in a 2 × 2 model. The metabolites — p-cresol, phenol, indole and ammonia — are produced as a result of fermentation of dietary protein in the gut by the resident microbiota and were quantified using SPME/GC-MS. Differences between dietary protein intake for p-cresol a, phenol b, indole c and ammonia d; sex differences in p-cresol d phenol e, indole f, and ammonia g; the interaction between dietary protein intake and sex for p-cresol i, phenol j, indole k and ammonia l were determined using the Kruskal Wallis test and Wilcoxon rank-sum pairwise comparisons with Benjamini-Hochberg correction. A significant threshold was set at adjusted p-value < 0.05
Urinary p-cresol concentrations were significantly higher in males than in females, regardless of their dietary protein intake (Fig. 7e) (p = 0.038). However, no differences were found in phenol (Fig. 7f), indole (Fig. 7g) or ammonia (Fig. 7h). Despite the elevated p-cresol levels in males, the impact of the HP diets on p-cresol was seen in females, not males. Only females on the HP diet exhibited significantly higher levels of urinary p-cresol compared to their counterparts on the SP diet (Fig. 7i) (p = 0.049), whereas males did not show significant differences in p-cresol levels between the two diets. The other metabolites produced by protein fermentation, phenol (Fig. 7j), indole (Fig. 7k), and ammonia (Fig. 7l), were stable across the sex by diet groups.
The impact of diet and sex on tight cell junction protein expressionThe expression of proteins located in TCJs in the intestinal epithelium was quantified using fluorescence immunohistology. Representational images of the barrier function-associated proteins, E-cadherin, a component of the adherens junction, and ZO-1 expression are shown in Fig. 8a. Consumption of a HP diet led to a significant reduction in the expression of E-cadherin in colonic tissue in comparison to the SP group (p < 0.001) (Fig. 8b), while no significant differences in ZO-1 expression were detected between the dietary groups (p = 0.16) (Fig. 8c). These results suggest that HP diets may impair gut barrier function by reducing E-cadherin expression, although they do not appear to affect intercellular ZO-1 when both males and females are analysed together. Similarly, no significant sex differences were observed for either E-cadherin (p = 0.15) (Fig. 8d) or ZO-1 expression (p = 0.89) (Fig. 8e). The expression of E-cadherin was significantly reduced in both females (p = 0.006) and males (p < 0.001) consuming the HP diet in comparison to the female and male SP groups (Fig. 8f). However, only females consuming HP diets showed a significantly lower ZO-1 expression (p = 0.044) than their same-sex SP group (Fig. 8g).
Fig. 8
Representational fluorescence immunohistology images of the colon in 56-day old female (F; n = 10) and male (M; n = 10) piglets fed either a standard protein (SP; 18% dietary protein) or a high protein (HP; 28% dietary protein) diet. Positive staining for E-cadherin (red) and zonula occludens-1 (ZO-1; green) a. The Log10 proportion of E-cadherin positive pixels b and ZO-1 c were quantified and compared between dietary groups; differences were also compared between males and females for E-cadherin d and ZO-1 e; the interaction between diet and sex was also compared for E-cadherin f and ZO-1 g. Ten individual images were analysed per piglets, with data presented as mean ± SEM and n = 5 piglets/sex/treatment group
Markers of inflammation and bacteria translocation across the epitheliumTo assess potential inflammation and intestinal permeability linked to high-protein diet-induced barrier dysfunction, IL-6 and LPS were measured in blood plasma from piglets. The change in IL-6 levels at day 56 from baseline showed no significant differences due to diet (Fig. 9a), sex (Fig. 9b), or any diet-by-sex interactions (Fig. 9c). Similarly, changes in plasma LPS concentrations did not significantly differ between diet groups (Fig. 9d), nor were there differences between females and males (Fig. 9e) or any diet-by sex interactions (Fig. 9f).
Fig. 9
Plasma samples from twenty 56-day-old piglets (n = 10 females, F; n = 10 males, M), fed either a standard protein (SP; 18% dietary protein) or high protein (HP; 28% dietary protein) diet for 28 days, were analysed for IL-6 and LPS concentrations using ELISA kits. The changes in systemic IL-6 concentration from baseline were calculated and compared for dietary protein groups a, sex b, and sex by protein group interactions c. Similarly, changes in systemic LPS were determined for protein groups d, sex e, and for sex by protein interactions f. Faecal microbiota from the piglets were sequenced using 16 S rRNA sequencing. Microbiota data were normalised using DESeq2, and Shapiro-Wilk normality tests were conducted on each variable. Based on the results of the Shapiro-Wilk test, either Pearson or Spearman correlation coefficients were calculated. Significant correlations after Benjamini-Hochberg correction are presented: females g, SP/M h, HP/M I. statistical significance was set at adjusted p-value < 0.05
Correlation analyses to determine relationships between microbial genera, p-cresol and TCJ proteinsTo determine the relationships between the gut microbiota and urinary metabolites (ammonia, indole, phenol, and p-cresol), IL-6, LPS, e-cadherin and ZO-1 protein expression, multiple correlation analyses were conducted. Depending on the distribution of the variables, either Pearson or Spearman’s correlation coefficient were used, with the Benjamini-Hochberg correction applied to account for multiple testing. When analysing all piglets together, no significant correlations were observed between any bacterial genera and the other variables. However, in subgroup analyses, a significant positive correlation was identified between Oscillospiraceae NK4A214 and plasma LPS levels in females (r = 0.9, p = 0.04) (Fig. 9g). Further subdividing the populations by both sex and dietary protein intake revealed a significant negative correlation between IL-6 and Lachnospiraceae_NK4A136 in the SP/M group (r = −0.1, p = 0.03) (Fig. 9h), as well as a significant negative correlation between Catenibacterium and LPS in the HP/M group (r = -0.99, p = 0.05) (Fig. 9i).
The influence of high protein diets on the expression of immune-associated proteins in the colonic mucosaDisruption in gut barrier function may influence immune activation within the lamina propria by enabling increased translocation of antigens through the epithelial barrier. To explore this potential relationship, the expression of immune-associated proteins in the colonic lamina propria of 56-day-old piglets was assessed using fluorescence immunohistology. Figure 10a depicts representative images of positive staining for CD45 (leukocytes), CD172a (Sirp-α, myeloid cells), MHC class II (antigen presenting cells), and capillary endothelium (MIL11). Co-expression was quantified to identify specific cell subsets and assess their potential functional roles within the LP. Representative images show co-expression of MIL11 (green), CD172a (red), and MHCII (blue), with CD172a+MILL11+ in yellow and CD172a+MHCII+ in magenta (Fig. 10b). Additionally, co-localisation of MHCII+ (green) and CD45+ (red) cells shows MHCII+CD45+ staining (yellow), perhaps indicating antigen-presenting cell activity (Fig. 10c).
Fig. 10
Fluorescence immunohistology representational images depicting the colon in 56-day old female (F; n = 10) and male (M; n = 10) piglets consuming diets composed of either standard protein (SP; 18% protein) or high protein (HP; 28% protein) a. Positive staining is shown for CD45, CD172a, MHCII, and MIL11. Representative images showing co-staining of MIL11, CD172a, and MHCII b as well as CD45 and MHCII c are also shown. The Log10 proportion of positive pixels for each region of interest (lamina propria) were quantified for each protein and compared between dietary protein groups d and sex e. The co-expression of CD45, CD172a, MHCII, and MIL11 proteins was determined between dietary protein groups and sex f as have the interactions between protein intake and sex g. The observed proportion of positive pixels for co-expression of CD172, MHCII, and MIL11 h and the combination of CD45, CD172a, MHCII and MIL11 i were compared against the expected counts of these combinations had the data had random distribution of expression of the proteins. Ten images were analysed per piglet and the data presented are mean ± SEM and n = 5 piglets/sex/treatment group
Differences in immune-associated protein expression between dietary protein groups, sexes, and interactions between sex and protein intake were quantified. High-protein diets, consumed for four weeks, resulted in significant reductions in the proportion of CD45+ cells in the colonic mucosa (log-transformed area; p = 0.006) when compared to the SP diet group. However, no significant differences were observed for CD172a+, MHCII+, or MIL11+ expression between dietary groups (Fig. 10d). Females exhibited significantly higher MHCII+ staining in the colonic mucosa compared to males (p < 0.001), while males had significantly more endothelial cell (MIL11+) staining relative to females (p = 0.01) (Fig. 10e).
To explore potential interactions between the target immune-associated target proteins, the overlap of positive staining for all four proteins were quantified (CD45+, CD172a+, MHCII+, and MIL11+), enabling the potential for antigen presentation between CD45+ leukocytes and MHCII+ monocytes and endothelial cells in the lamina propria to be assessed. When comparing these interactions across dietary groups, no significant differences were found. However, a trend was observed, perhaps indicating higher levels of antigen presentation in males compared to females (p = 0.071) (Fig. 10f) which may have become significant if more animals had been used in the study. Lastly, no significant interactions between sex and dietary protein groups were detected in the expression of CD45, CD172a, MHCII, or MIL11 (data not shown). Nonetheless, a trend towards increased antigen presentation by endothelial cells was noted in the high-protein male group (HP/M) compared to the high-protein female group (HP/F) (p = 0.057) (Fig. 10g). Evidence is provided to demonstrate that the co-expression of CD172a, MIL11, MHCII and CD45 observed was significantly higher than that expected from random distribution (p < 0.01) (Fig. 10h), whereas when CD45 was omitted (Fig. 10i), the amount of co-staining observed was significantly lower than expected from random distribution, given the amount of single staining measured (p < 0.01) for all treatment and sex combinations explored. No other differences were found in the co-expression of immune cells associated proteins; therefore data is not shown.
Comments (0)