
Remember me
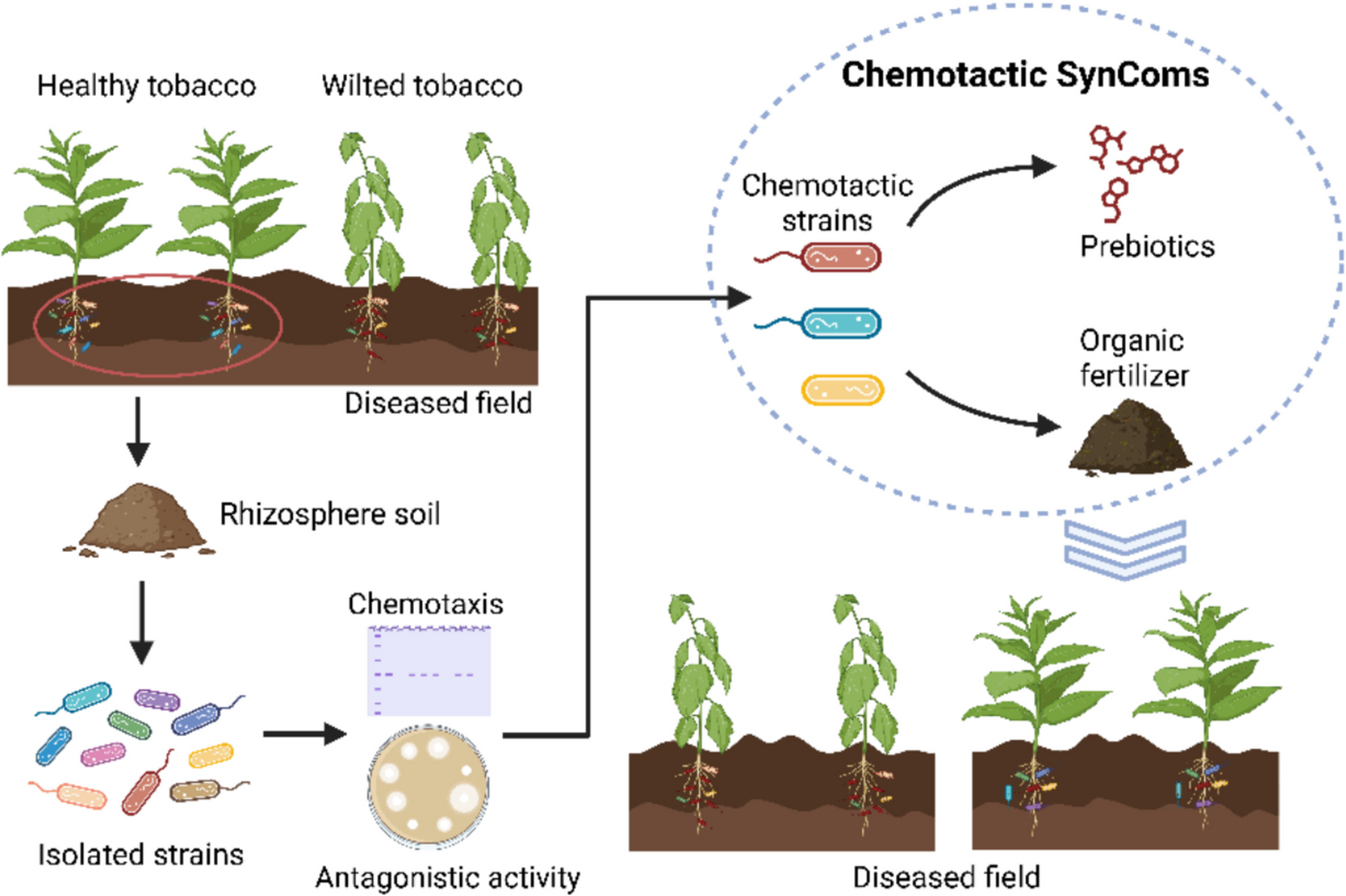
In previous work on the genetic engineering of the strictly anaerobic cellulose-degrading bacterium C. cellulovorans, we reported an efficiency of transconjugation from E. coli donor cells of about 10−8–10−7 transconjugants per recipient cell for the C. cellulovorans 743B wildtype strain (Schöllkopf et al. 2025). Applying in vivo methylation in E. coli with a C. cellulovorans methylase to protect the transconjugated DNA from restriction by the type II restriction endonuclease present in C. cellulovorans enables the transfer of recombinant plasmids, but the conjugation efficiency remained poor (Fig. 1a). Interestingly, the genetic accessibility was greatly improved in a strain that arose spontaneously during genetic engineering experiments, where we aimed to delete the engE gene to study the role of EngE, a cellulosomal endoglucanase, for cell surface attachment of the cellulosome. The strain was designated as C. cellulovorans ΔengE/2. The conjugation efficiency with this strain as recipient was approximately 10−3 transconjugants per recipient cell (Fig. 1b) compared to about 10−7 for the wildtype. Thus, we sequenced the chromosomal DNA of the mutant using Nanopore technology to determine potential genetic changes that could be responsible for the strain’s improved ability for plasmid uptake by conjugation. Three major chromosomal deletions were identified in strain ΔengE/2 (Fig. 1c–f): Deletion A (~ 129 kb), which ranged from Clocel_3950 to Clocel_4071, included the type II RM system operon that we have characterized before by generating a type II restriction deficient strain. The targeted deletion of this type II RM system resulted in a transconjugation efficiency of approximately 10−6 transconjugants per recipient cell (Schöllkopf et al. 2025). In a second deletion region (deletion B, ~ 17 kb), ranging from Clocel_2327 to Clocel_2351, a devR/cas7 ortholog (Clocel_2327) and a cas8 ortholog (Clocel_2328) of a putative endogenous CRISPR-Cas system were no longer present. The third deletion (deletion C, ~ 23 kb) involved the genes Clocel_0831 to Clocel_0854, including genes belonging to a putative Wadjet operon.
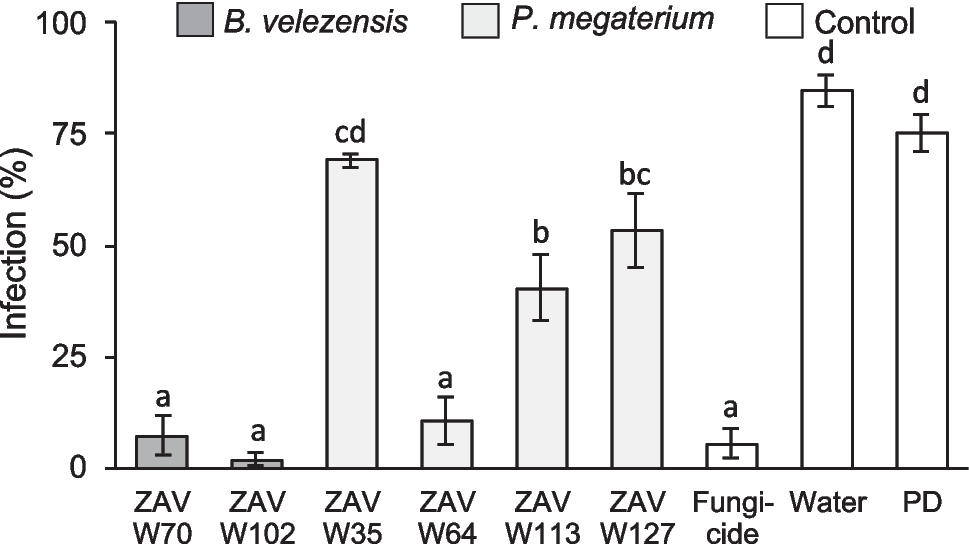
Fig. 1
Improvement of transconjugation efficiency in a spontaneous mutant of C. cellulovorans. a Selection plate with merely few C. cellulovorans transconjugant colonies (marked with arrows) after a representative mating experiment with E. coli NEB10B-pMTL85131/placORMI as donor and the C. cellulovorans 743B wildtype strain as recipient. b Selection plate after mating with E. coli NEB10B-pMTL85131/placORMI as donor and the C. cellulovorans mutant strain ΔengE/2 as recipient. c Comparison of the genome sequence of C. cellulovorans 743B WT and the mutant strain ΔengE/2 by dot plot analysis (Cabanettes and Klopp 2018). The three deletion regions A, B and C that were found to be missing in the chromosome of the ΔengE/2 mutant strain are marked with orange ellipses. d Chromosomal location of deletion A. e Chromosomal location of deletion B. f Chromosomal location of deletion C
Deletion of the putative devR and cas8 genes readily occurs spontaneouslyThe putative CRISPR locus of C. cellulovorans contains a CRISPR array of 35 spacers, Clocel_2325 (49.27% similarity to a CRISPR-associated helicase Cas3 of Anoxybacillus tepidamans), Clocel_2326 (55.26% similarity to a hypothetical protein in Clostridium butyricum, and 39.56% to a type I-A Cas5a in Bacillus thuringiensis), Clocel_2327 (64.09% similarity to DevR of C. butyricum), and Clocel_2328 (34.18% similarity to a hypothetical protein of C. butyricum). Genome annotation of this region of the C. cellulovorans chromosome indicated cas3, cas5, devR/cas7 and cas8b/cas8a2 genes, representing part of a putative type I-B CRISPR system (Schwengers et al. 2021; Alkhnbashi et al. 2021).
Analysis of the cas ORFs, coding for the CRISPR locus indicated the spontaneous deletion of the putative cas ORFs in various independent cultures of C. cellulovorans. To confirm the nucleotide sequence of the chromosomal region carrying the cas ORFs in the wildtype strain, a glycerol stock from an early culture frozen away after the strain had been obtained from the DSMZ culture collection in 2020 was reactivated and inoculated into DMSZ 520 medium with 5 g L−1 cellobiose at 37 °C and the integrity of the chromosomal cas region was monitored over time. The cas region’s presence was confirmed in early cultures by PCR, but the region was spontaneously lost after around six consecutive sub-cultivations (Fig. 2).
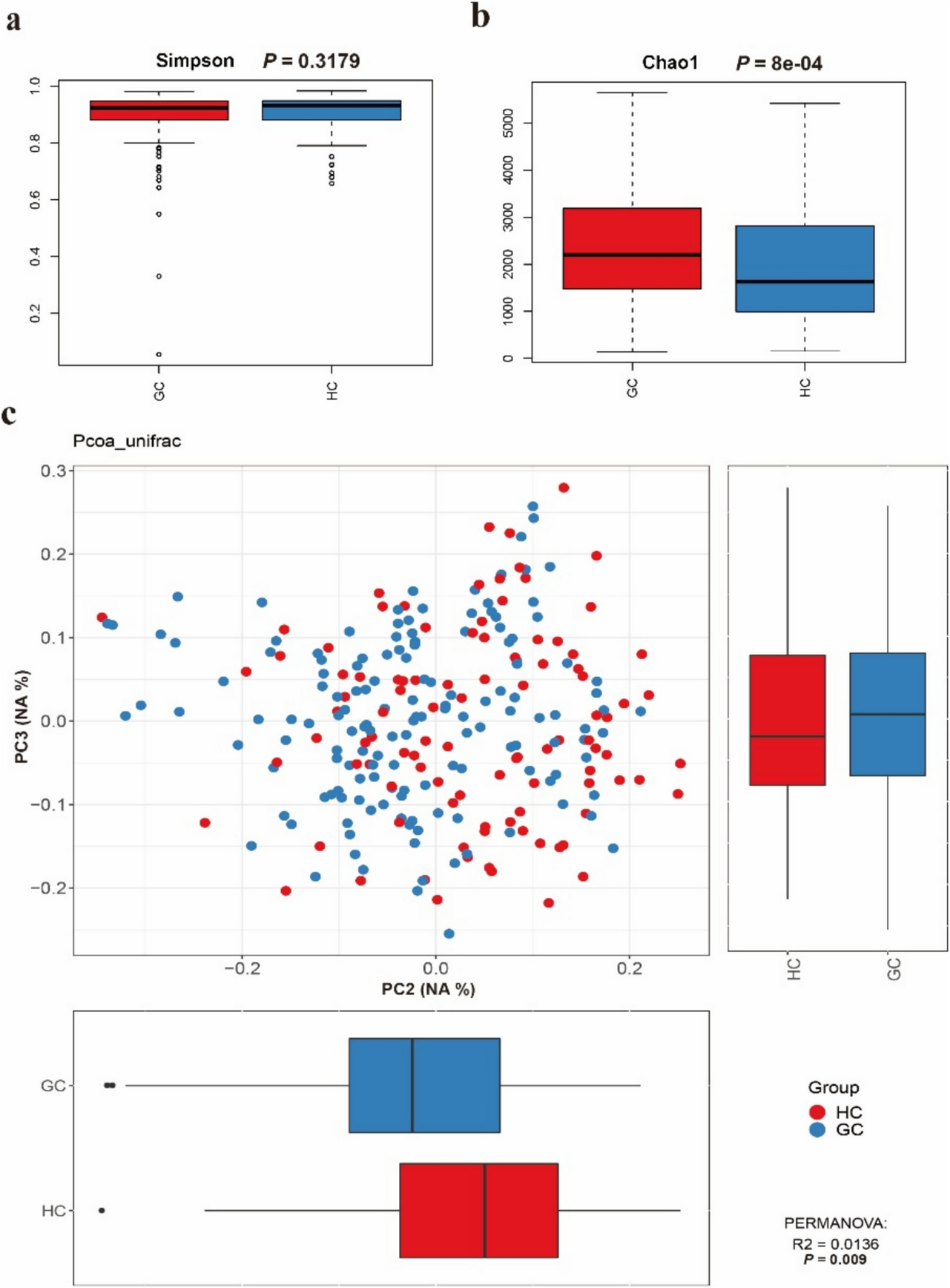
Fig. 2
Spontaneous deletion of the cas locus. a The gDNA of wildtype C. cellulovorans 743B was isolated after 3 to 17 consecutive sub-cultivations after the strain was obtained from DSMZ (P3, P4, P6, P7, P8, P11, and P17). The cas locus was amplified using primers D5 and D6, resulting in a 6 kb product, while no product was expected when the locus was spontaneously deleted (lanes 1–15, the PCR was performed in technical duplicates). The integrity of the gDNA was controlled using primers D7 and D8 to amplify the Clocel_4007-4008 locus (lanes 16–22). b C. cellulovorans 743B chromosomal cas locus. The identity of the ORF cas8 is hypothetical and requires further confirmation. The binding sites of the check primers Del5 and Del6 are indicated with arrows. The confirmed deletion of C. cellulovorans ΔengE/2 (see Fig. 1e) is shown in grey
To determine if these spontaneous cas deletion mutations appearing after repeated sub-cultivation were the same as observed in the three-fold deletion strain ΔengE/2, genome resequencing of four cultures, which were inoculated from different glycerol stocks, was carried out and identified deletions in the Clocel_2327 gene downstream of the sequence 5′-…TTGCC ACTAT CACTT A-3′ and upstream of 5′-ACTTA TATGT AAAGT T…−3′ in Clocel_2352. Apparently, a fusion of the “ACTTA” sequence motif which is present in both the Clocel_2327 and the Clocel_2352 genes had occurred.
The CRISPR spacer sequences were investigated by BLASTn similarity search against the core_nt database to identify potential targets in plasmids or invasive DNA elements like phages. However, no potential target of the CRISPR spacers was identified; thus, their origin and the functionality of the C. cellulovorans 743B CRISPR-Cas system remain obscure.
Similarity of the Jet proteins of C. cellulovorans with the Jet proteins of Bacillus cereus and other putative Jet protein orthologsPrevious work led to discovery of the Wadjet system of Bacillus cereus Q1 by introduction of the corresponding genes comprising this system into a heterologous host, B. subtilis (Doron et al. 2018). Comparative sequence analysis of the putative C. cellulovorans Wadjet proteins encoded by ORFs Clocel_0831 to Clocel_0834 revealed about 25.16%, 26.53%, 35.68%, and 26.62% similarity with Bacillus cereus Q1 JetA, JetB, JetC, and JetD, respectively. Next, the putative nuclease JetD, which is the signature protein of the Wadjet operon, was analyzed in detail. Previous reports revealed four conserved and catalytic active amino acid residues in JetD from Bacillus cereus Q1 and Pseudomonas aeruginosa PA14 (Deep et al. 2022). We carried out an amino sequence alignment of the putative JetD proteins of C. cellulovorans 743B, Clostridium beijerincki DJ123 and Clostridium kluyveri JZZ to identify conserved residues. C. beijerincki DJ123 served as an example for clostridia without a reported plasmid, while C. kluyveri JZZ carries a 58.6 kb plasmid. The alignment shown in Fig. 3 confirms the conservation of the essential amino acid residues in all JetD primary structures. This observation reinforces the assumption of a functional Wadjet system in C. cellulovorans. An AlphaFold prediction of the three-dimensional fold and positions of the active site residues of C. cellulovorans JetD, as well as a structural comparison with other predicted or experimentally confirmed JetD structures, is shown in supplemental Figure S2.
Fig. 3
Alignment of JetD amino acid sequence segments. The protein sequences of JetD proteins from Bacillus cereus Q1, Pseudomonas aeruginosa PA14, C. cellulovorans 734B, Clostridium kluyveri JZZ, and Clostridium beijerincki DJ123 were aligned using Jalview’s Muscle algorithm with default settings. Conserved amino acids are shown in blue, and the experimentally confirmed essential residues of the JetD enzymes from Bacillus cereus Q1 and Pseudomonas aeruginosa PA14 are indicated with arrows
Deletion of the chromosomal jetABCD genes in C. cellulovorans 743B and its effect on transconjugation efficiencyThe functionality of the putative Wadjet system of C. cellulovorans was experimentally evaluated by markerless deletion of the jetABCD genes from the C. cellulovorans chromosome. Flanking regions, covering 1 kb each up- and downstream of the putative Wadjet system-encoding genes (jetABCD, ORFs Clocel_0831, Clocel_0832, Clocel_0833, and Clocel_0834), were designed to retain the start of Clocel_0831 and stop codon of Clocel_0834. The fused flanks were inserted into the vector pNick, and the seed sequence of the guide RNA was designed to target the Clocel_0834 (jetD) locus.
After transconjugation, two colonies obtained were cultivated overnight in selection medium, and subsequently spread onto induction plates containing 10 µg mL−1 thiamphenicol for plasmid maintenance and 5 mM theophylline for induction of the cas9 nickase. After incubation, eight colonies appeared, all with wildtype jet genotype according to PCR analysis. Subsequently, these colonies were re-streaked onto induction plates containing thiamphenicol and theophylline, and one clone with a mixed genotype (as determined with PCR using primer pairs resulting in products of different length for the wildtype and the anticipated jet mutant genotypes) was obtained. A third round of re-streaking onto induction plates finally resulted in clones with the desired deletion genotype. Further analysis by PCR confirmed the chromosomal deletion of the ORFs Clocel_0831 (jetA), Clocel_0832 (jetB), Clocel_0833 (jetC), Clocel_0834 (jetD), which resulted in a 4.3 kb long PCR product for the clean deletion mutant in comparison to the 10.7 kb long fragment for the wildtype. Curing of the deletion plasmid was successful after repeating five times in succession inoculation and growth in medium without antibiotic pressure (Fig. 4). Compared with the wildtype, the growth behavior and cell morphology of the ΔjetABCD strain were not affected. Lastly, the deletion was confirmed by Illumina whole-genome sequencing.
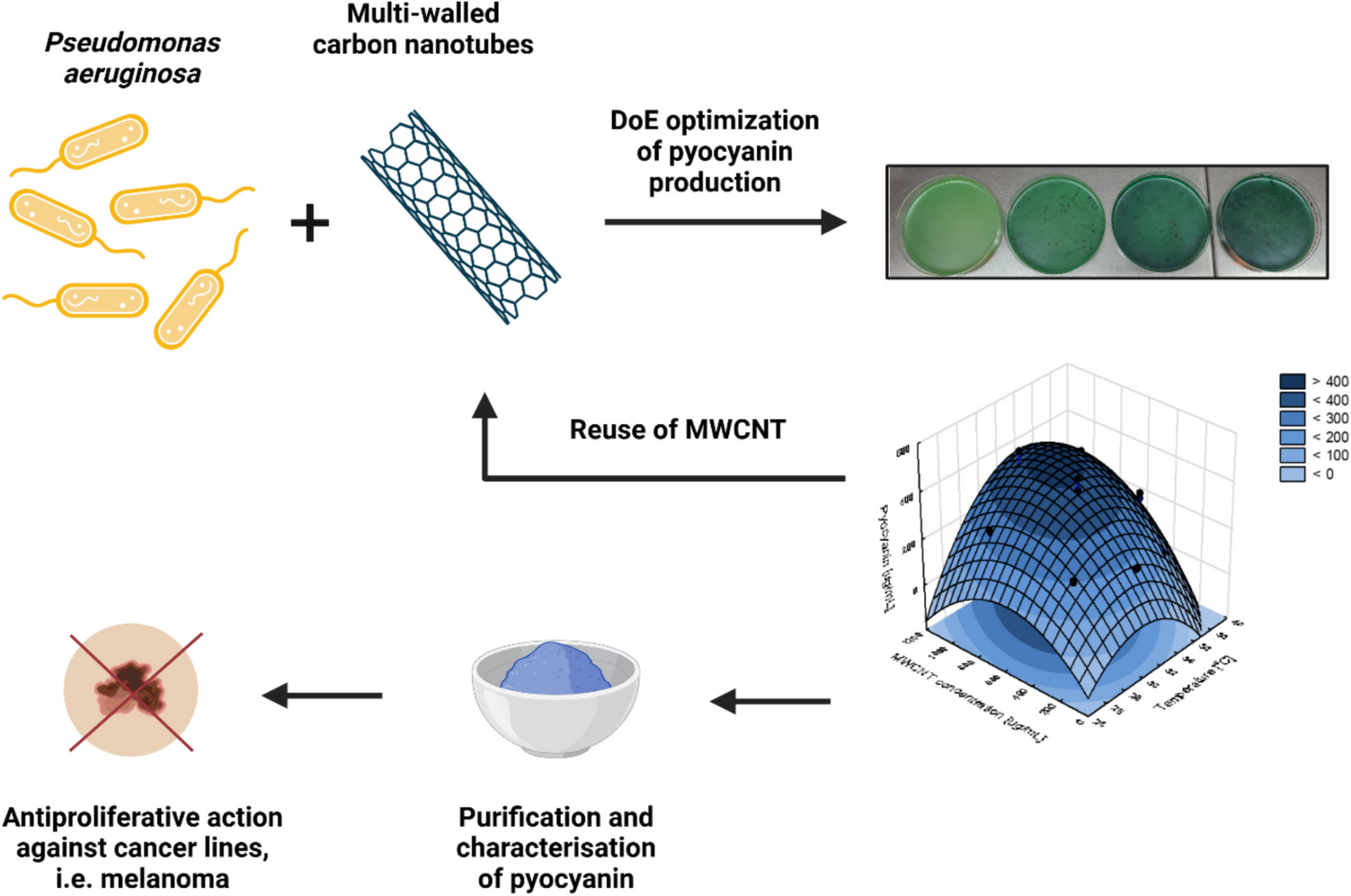
Fig. 4
Deletion of the putative Wadjet gene cluster jetABCD in the C. cellulovorans 743B wildtype strain. a PCR analysis with primers D1 and D2 either confirmed the chromosomal deletion of jetABCD (4.3 kb) or showed the wildtype genotype (10.7 kb) (lanes 1–5). Confirmation of plasmid curing was done by PCR with primers D3 and D4 targeting cas9 (4.1 kb) (lanes 6–10). Lanes 1 and 6, C. cellulovorans ΔjetABCD K1; lanes 2 and 7, C. cellulovorans ΔjetABCD K2; lanes 3 and 8, C. cellulovorans ΔjetABCD K4; lanes 4 and 9, C. cellulovorans 743B wildtype; lanes 5 and 10, pNick_delJetABCD-sg1 plasmid. b C. cellulovorans 743B chromosomal Wadjet locus. The check primers Del1 and Del2 are indicated with arrows. The confirmed deletion of C. cellulovorans ΔjetABCD is shown in black
The function of the Wadjet system as barrier against incoming plasmid DNA in C. cellulovorans was investigated by evaluation of the impact of the jetABCD deletion on transconjugation of a recombinant plasmid from E. coli donor cells. We have previously shown that the transconjugation efficiency was substantially improved by in vivo methylation in E. coli of the shuttle plasmids via expression of a C. cellulovorans-born methyltransferase from a type II RM system (Schöllkopf et al. 2025). Therefore, we tested transconjugation of the un-methylated and the in vivo methylated pMTL83151 plasmid (4.5 kb) from the respective E. coli donor strains, E. coli NEB10B-pMTL85131, or E. coli NEB10B-pMTL85131/placORMI, respectively (Fig. 5a and b). A transfer frequency of 2.42 ± 0.94 × 10−3 transconjugants per ΔjetABCD recipient cell was obtained without previous in vivo methylation (donor strain E. coli NEB10B-pMTL85131), corresponding to an increase by approximately five orders of magnitude compared to conjugation of the same plasmid from the E. coli donor to wildtype C. cellulovorans 743B recipient cells. By employing in vivo methylation (donor strain E. coli NEB10B-pMTL85131/placORMI), the transconjugation frequency was further increased to 1.71 ± 0.27 × 10−2 transconjugants per ΔjetABCD recipient cell (n = 5, mean ± SD) (Fig. 5c).
Fig. 5
Conjugation efficiency of plasmid pMTL83151 into C. cellulovorans ΔjetABCD. Panels a and b show selection plates after mating of pMTL83151-bearing E. coli donor strains with C. cellulovorans ΔjetABCD as recipient. a Mating experiment using E. coli NEB10B-pMTL85131 (unmethylated plasmid) as donor and C. cellulovorans ΔjetABCD as recipient. b Mating experiments were performed using E. coli NEB10B-pMTL85131/placORMI (plasmid methylated in vivo via expression of a C. cellulovorans-born methyltransferase of a type II RM system) as donor and C. cellulovorans ΔjetABCD as recipient. c Comparison of the conjugation efficiency of different C. cellulovorans strains using unmethylated plasmid (black bar) and methylated plasmid (striped bar). The efficiency values are given as number of transconjugants per recipient cell with standard deviation of five independent experiments for ΔjetABCD (n = 5, mean ± SD). *Data for WT and ΔTypeII are taken from Schöllkopf et al. (2025). The experimental conditions (see Materials and Methods) were the same in all experiments
Wadjet systems in other BacillotaSince Wadjet systems act against the presence of plasmid DNA, the distribution of putative Wadjet system-encoding genes and at the same time the occurrence of natural plasmids in the phylum Bacillota (formerly Firmicutes) was analyzed. For this purpose, the coding sequences of the JetA, JetB, JetC, and JetD proteins of Bacillus cereus Q1 were blasted with tBLASTn against the “RefSeq Genome Database (refseq_genomes)” including “Bacillus/Clostridium group (taxid: 1239),” without “Models” or “uncultured/environmental sample sequences” (data acquisition: August 2024).
As mentioned before, jetD of Bacillus cereus Q1 encodes a nuclease essential for the Wadjet-catalyzed plasmid cleavage, so further work was focused on this protein. About 1,442 strains were identified to putatively possess orthologues of jetD, while plasmids were identified in only 32 (2.2%) of these strains (supplementary data S3 to S5). Among these plasmid-bearing strains, the plasmid sizes were between 500 kb and 50 kb, i. e. above the suggested Wadjet threshold size of 50–100 kb (Liu et al. 2023), in 16 cases; five Enterocloster bolteae strains with a > 40 kb plasmid also carried jetABCD genes. Nine of the strains potentially bear plasmids of < 50 kb but lack at least one jet gene ortholog. The co-occurrence of the four jet genes and plasmids is visualized in Fig. 6, revealing 17 strains bearing jetABCD and a plasmid, which are listed in Table S3. Selenomonas sp. oral taxon 136 F0591 (NZ_CP014239.1) and Anaerostipes sp. PC18 (NZ_CP121164.1) appear to carry relatively small plasmids of 15.6 kb and 1.3 kb, as well as putative jetABCD genes. However, in these cases the sequence data was retrieved from metagenomic samples of supragingival dental plaque of a molar tooth, and the intestinal contents of Mus musculus. Because these genome sequences were not obtained from axenic clonal isolates, uncertainty remains if these uncultivated strains indeed simultaneously carry chromosomal jetABCD genes and small-sized plasmids.
Fig. 6
Co-occurrence of jetA, jetB, jetC orthologs and plasmid/jetD ortholog-bearing Bacillota strains with sequenced genomes
Comments (0)