
Remember me
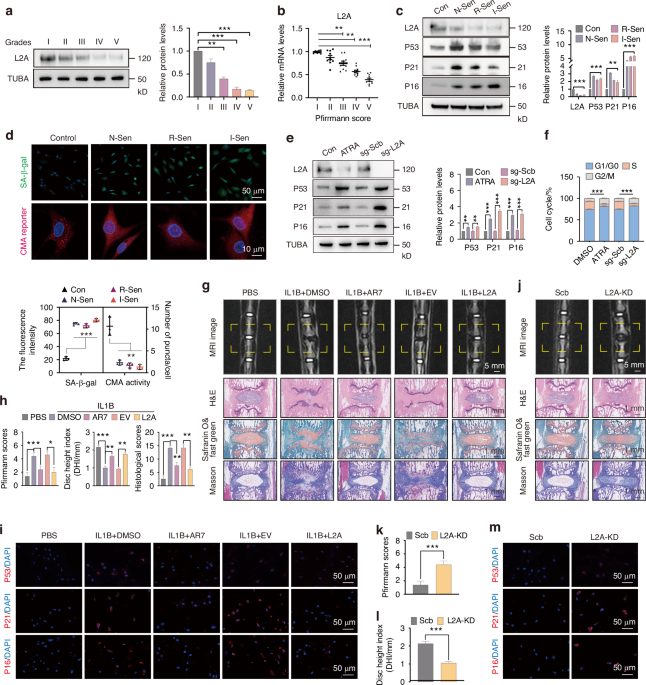

To investigate the molecular etiology of age-related skeletal degeneration, we performed single-cell RNA sequencing (scRNAseq) of the hindlimb skeletal elements of young adult (3-month-old) and middle-aged (12-month-old) mice (Fig. 1a, Fig. S1a, b). We analyzed middle-aged mice (10–14 months) rather than aged (18–24 months) mice to identify factors involved in the progression of skeletal aging rather than in the end product, the irreversibly aged skeleton. Uniform manifold approximation and projection (UMAP) visualization distinguished 24 cell clusters, which we classified as 22 hematopoietic and endothelial and 2 stromal/osteolineage populations based on published scRNAseq data sets23,24 (Fig. 1b). Of the 2 stromal clusters, cluster 22 expressed genes characteristic of Lepr+ SSPCs, including Lepr, Cxcl12, and Kitl15 (Table S1). Published scRNAseq studies show that Lepr+ SSPCs exist in two states: osteo-primed and adipo-primed.25 With aging, there is an expansion of the adipo-primed population together with an increase in adipocyte-associated gene expression.7,26 We similarly observed an adipogenic shift in the middle-aged Lepr+ SSPC cluster with increased adipogenic gene expression (Apoe, Lpl, Cebpa/b, Adipoq), increased expression of anti-osteogenic BMP inhibitors, and a decrease in pro-osteogenic pathways such as Bmp4 and Egfr5 (Fig. S1c).
Fig. 1
Aging activates Notch signaling genes in skeletal stem cells. a Schematic of experimental workflow for scRNAseq of young adult (~3-month-old) and middle-aged (~12-month-old) bone stromal (CD31−CD45−TER119−) and hematopoietic and endothelial compartments (CD31+CD45+TER119−) from hindlimb bone tissue sorted and combined at a 1:1 ratio, then processed for scRNAseq. b scRNAseq UMAP-based visualization of stromal, hematopoietic, and endothelial bone and bone marrow populations with 24 clusters. Young n = 15 225 cells, Middle-aged n = 8 831 cells. The pink outline indicates the stromal compartment and the green outline indicates the hematopoietic and endothelial compartment. c Gene set enrichment analysis (GSEA) using the package escape 1.4.1. GSEA analysis was performed on the stromal hematopoietic, and endothelial compartments separately. Split Violin plot shows normalized enrichment score (NES) for hallmark_notch_signaling from the H:hallmark gene sets between young and middle-aged stromal (FDR = 1.830 372e−07, young median NES = 536.031, middle-aged median NES = 927.393), hematopoietic (FDR = 1.032 392e-02, young median NES = 939.566, middle-aged median NES = 899.742), and endothelial populations (FDR = 1.000 000, young median NES = 695.455, middle-aged median NES = 437.276) Wilcoxon test. d Dotplot showing expression of Notch signaling pathway genes in cluster 22, which is mainly comprised of Lepr+ expressing SSPCs between middle-aged and young mice. e SSPCs were subjected to ATAC sequencing. Compared to young SSPCs, chromatin in middle-aged SSPCs was more accessible around the transcriptional start site (TSS) in genes associated with Notch signaling
To explore the pathways responsible for the adipogenic shift with age, we used gene set enrichment analysis (GSEA) to define transcriptional signatures that were enriched in the stromal and osteolineage populations during aging and thus could be involved in age-related skeletal degeneration (Fig. S1d). Evidence from human, young and middle-aged bone samples suggests Notch signaling components to be elevated with aging.27 Utilizing the MsigDB “hallmark” gene set,28 NOTCH_SIGNALING exhibited a significant increase with aging in stromal/osteo-lineage cells (FDR = 1.830373e−07) (Fig. 1c). We confirmed this observation by qRT-PCR, finding that the expression of key Notch pathway components is elevated within middle-aged bones (Fig. S1e). In contrast, the GSEA NOTCH_SIGNALING did not show a difference in the hematopoietic or endothelial populations (Fig. 1c), although the Notch ligand Delta4, Dll4, was upregulated in bone marrow endothelial cells with aging, suggesting an increase in Notch signaling activity (Fig. S1f). To interrogate the stromal and osteolineage compartments at higher resolution, we repeated UMAP clustering without hematopoietic and endothelial cells, discerning 5 clusters representing 3 distinct populations (Fig. S2a, b). Cluster 4 was SSPC-like, defined by Cxcl12, Lepr, and Kitl15 (Fig. S2c). Clusters 0–2 resembled more committed osteoprogenitors, expressing Ly6a (Sca-1), Thy1,29Mfap5,7Itga5,30 and Postn31 (Fig. S2d). Cluster 3 was enriched for markers of terminally differentiated osteoblasts, such as Col1a1, Bglap, Gsn, Clec3b, and chondrocytes, such as Comp and Acan (Fig. S2e). The osteoprogenitor (clusters 0–2) and osteo/chondro mature lineage cells (cluster 3) were considerably depleted during aging (Fig. S2b), consistent with the age-related loss of bone mineral density. These analyses also showed that Notch pathway components and downstream targets were elevated primarily in Lepr+ SSPCs with age (cluster 4) (Fig. 1d and Fig. S3a), consistent with previous human and mouse studies.27,32
Aberrant gene expression during aging has been linked to a loss of repressive heterochromatin.33,34 To determine whether the transcriptional changes in SSPCs during aging were associated with a transformation of the epigenetic landscape, we performed ATAC-sequencing on young and middle-aged SSPCs. In line with previous observations, middle-aged mice showed a global increase in chromatin accessibility with aging (Fig. S3b). Moreover, chromatin surrounding the transcriptional start site (TSS) of Notch-associated loci became more accessible with age (Fig. 1e), consistent with the observed increase in transcription. Thus, age-related skeletal degeneration is accompanied by epigenetic and transcriptional dysregulation of Notch pathway components in Lepr+ SSPCs.
Loss of Notch signaling in SSPCs promotes transcriptional osteo-primingInhibition of Notch signaling promotes osteogenesis and reduces adipogenesis in vitro.32 However, it is unknown whether Notch regulates the detrimental shift from osteogenesis to adipogenesis in the aging skeleton. To investigate this, we disrupted Notch signaling activity in SSPCs in vivo. We focused on Lepr+ SSPCs since they are the main source of osteoblasts and adipocytes in adulthood15,35 and exhibit dysregulated Notch-associated gene expression during aging (Fig. 1d). Nicastrin (Ncstn) is a γ-secretase that activates Notch signaling by cleaving all four Notch receptors. Consequently, loss of Ncstn abolishes Notch signaling.36 We combined Ncstnfl/fl mice36 with a Lepr Cre driver,37 and performed scRNAseq of middle-aged Ncstnfl/fl (control) and LeprCre; Ncstnfl/fl (Ncstn cKO) femurs and tibiae (Fig. 2a), enriching for skeletal and stromal lineages, as in Fig. 1a. Notch pathway genes were significantly downregulated in Ncstn cKO SSPCs compared to control cells (Fig. S4a, b) confirming that Ncstn deletion impaired Notch signaling. We also validated this by qRT-PCR for the Notch downstream targets Hey1 and Hes118,38,39 (Fig. S4c), UMAP visualization distinguished 25 hematopoietic and endothelial and 1 stromal cluster (Fig. 2a). Analysis of the stromal population in isolation revealed 4 clusters (Fig. 2b) that we defined as Lepr+ SSPCs (cluster 1, expressing Lepr and Cxcl12), early osteolineage progenitor cells (Early OLCs) (cluster 2, expressing Pdgfra, Ly6a, Cd34, and Mfap5), differentiated osteolineage cells (Late OLCs) (cluster 0, expressing Col1a1), and bone marrow endothelial cells (BMECs) (cluster 3, expressing Cdh5) (Fig. 2c, d). The stromal compartment from Ncstn cKO mice had a reduced proportion of SSPCs and an increased proportion of early and late osteoprogenitors compared to controls (Fig. 2e). We confirmed these data by flow cytometry, observing a decrease in the frequency of LEPR+ SSPCs (Fig. 2f) and an increase in PDGFRα+ and SCA-1+ osteoprogenitor frequency in Ncstn cKO mice (Fig. 2f, Fig. S5).
Fig. 2
Decreased Notch signaling during aging increases osteogenic and decreases adipogenic gene signatures. a UMAP clusters from control Ncstnfl/fl (n = 8 831 cells) and LeprCre; Ncstnfl/fl cKO (n = 9 439 cells) middle-aged digested bone and bone marrow showing a stromal (outlined in pink) and hematopoietic/ endothelial compartment (outlined in green). b UMAP after sub-clustering of the stromal compartment of control and Ncstn cKO middle-aged mice. SSPC skeletal stem and progenitor cell, OLC osteo-lineage cell, BMEC Bone marrow endothelial cell. c Heatmap showing the separation of the 4 clusters. d Violin plot showing expression of marker genes for each cluster and of SSPC (Lepr, Cxcl12), osteolineage cells (Cd34, Ly6a, Pdgfra, Mfap5, Col1a1), and bone marrow endothelial cells (Cdh5). e Proportion of each cluster found in control and Ncstn cKO mice. f Flow cytometry analysis to confirm the proportional shift seen in scRNAseq of primitive Lepr+ SSPCs and more mature osteolineage populations PDGFRα+ and SCA-1+ cells between middle-aged control (n = 5) and Ncstn cKO (n = 6) mice. *P < 0.05, **P < 0.01, ***P < 0.001. Data were represented as mean ± s.e.m
To understand the basis of this change, we focused on the Notch signaling deficient SSPC population. We identified 262 genes that were upregulated and 313 that were downregulated in Ncstn cKO vs. control Lepr+ SSPCs (cluster 1, P < 0.05) (Fig. 3a, Table S2). Upregulated genes included the osteogenic markers Bglap2, Myoc,40Ncam1,41Col1a1, Wnt4a,42Clec11a,43,44Foxp145 (Fig. 3a, b). Conversely, downregulated genes included adipogenic-associated factors, such as Cebpa, Cebpb,46Junb,47Ccl2,46,47,48,49 and inhibitors of osteogenesis, Socs350 and Grem15 (Fig. 3a, b). Thus, reducing Notch signaling activity in SSPCs results in elevated osteolineage gene expression, which may transcriptionally “prime” cells for osteogenic differentiation. To examine progenitor number, self-renewal, and osteo-priming at a functional level, we performed colony-forming unit (CFU-F) assays, whereby each stem and progenitor cell gives rise to an individual colony. Bone marrow cells from Ncstn cKO mice gave rise to more and larger CFU-Fs than controls (Fig. 3c, d), signifying a greater number of progenitors with increased self-renewal capacity. Notably, CFU-Fs from Ncstn cKO mice also displayed elevated alkaline phosphatase (ALP) enzymatic activity, one of the earliest osteogenic markers (Fig. 3e, f), indicating that progenitors are functionally as well as transcriptionally more osteo-primed than those from control mice. The proportion of colonies exhibiting ALP activity was significantly reduced upon overexpression of Hes1 (Fig. 3g, h), demonstrating that Ncstn cKO SSPC osteo-priming was a direct effect of decreased Notch signaling.
Fig. 3
Loss of Notch signaling during aging increases the number of osteoprogenitors. a Volcano plot showing differential gene expression between Ncstn cKO and control SSPC cluster. Red genes are upregulated in Ncstn cKO SSPCs and blue genes are downregulated in Ncstn cKO SSPCs compared to control (average log2FC > 0, P value < 0.05). b Violin plot showing expression of adipogenic, anti-osteogenic, and osteogenic genes between control and Ncstn cKO SSPCs. c Colony-forming unit fibroblast assay (CFU-F) of control and Ncstn cKO middle-aged bone marrow showing representative colony staining with crystal violet. d Quantification of CFU-F efficiency (number of cells plated/number of colonies formed) and colony diameter (in pixels)(n = 3). e Colony-forming unit fibroblast assay (CFU-F) of control and Ncstn cKO middle-aged bone marrow showing representative colony staining for alkaline phosphatase (ALP) enzymatic activity, an early osteogenic differentiation marker. f Quantification of CFU-Fs showing specified percentages of ALP+ cells per colonies (n = 3). g Colony-forming unit fibroblast assay (CFU-F) of Ncstn cKO middle-aged bone marrow showing representative colony staining with ALP with a control plasmid or Hes1 over-expression plasmid. High-magnification image of a representative colony. h Quantification of CFU-Fs showing specified percentages of ALP+ cells per colonies (n = 3). *P < 0.05, **P < 0.01, ****P < 0.000 1. Data were represented as mean ± s.e.m
Since aging is typically associated with a decrease in osteo-primed and an increase in adipo-primed SSPCs,7,26 we then asked whether the stromal compartment of Ncstn cKO middle-aged mice more closely resembled that of young rather than middle-aged control mice. To do so, we integrated our young and middle-aged control, and middle-aged Ncstn cKO scRNAseq datasets, isolated the stromal population and performed sub-clustering (Fig. S6a). We identified several clusters that expressed periosteal genes, a cluster with an SSPC transcriptional signature, and a cluster expressing bone marrow osteo-chondro lineage markers including chondrocyte, early OLC, and late OLC markers (Fig. S6b–f). Notably, aging in control mice was associated with a decline in bone marrow OLCs that was averted in aging cKO mice (Fig. S6a, g). In contrast, the periosteal clusters were greatly diminished in both middle-aged control and cKO mice (Fig. S6a). The specific rescue of bone marrow OLCs in aging LeprCre; Ncstnfl/fl (cKO) mice is consistent with the fact that Lepr+ SSPCs predominantly give rise to trabecular bone, not cells within the periosteum.15 Together, these data demonstrate that moderating Notch signaling in Lepr+ SSPCs prevents the loss of osteo-priming and reduction in bone marrow osteoprogenitors with age.
Ncstn cKO mice exhibit increased bone mass and reduced bone marrow adiposityNext, we examined how these transcriptional and cell population-based changes in Ncstn cKO mice affected the skeletal aging phenotype in vivo. While Ncstn cKO mice displayed no gross phenotypic differences compared to littermate controls (Fig. S7a, b), microcomputed tomography (microCT) analysis of middle-aged cKO femurs showed a substantial increase in trabecular bone mass throughout the bone marrow cavity compared to controls (Fig. 4a). Specifically, we detected a significant increase in BV/TV, trabecular thickness (Tb.Th) and trabecular number (Tb.N) and a decrease in trabecular spacing (Tb.Sp) (Fig. 4b). No difference was observed in cortical bone parameters (Fig. S7c). The high bone mass phenotype was also observed in other long bones (data not shown) and, in the vertebral column (Fig. S7d), where it correlated with improved mechanical properties of the cancellous bone (Fig. S7e). The increase in trabecular bone persisted in aged 2-year-old mice (Fig. S7h, i). The phenotype was sex- and age-dependent with the increase in bone mass more pronounced in femurs from female mice (Fig. 4c, d), even at a younger age (~12-week-old) (Fig. S7f, g). Female Ncstn cKO middle-aged mice also showed an increase in mineral apposition rate compared to middle-aged female control mice (Fig. 4f) Lineage tracing, using a Ncstn cKO; tdTomato mouse, established that the trabecular bone within the marrow cavity was derived directly from Lepr+ SSPCs (Fig. 4e). To determine whether Ncstn cKO mice showed a reduction in catabolic bone resorption, that could additionally contribute to the high bone mass phenotype, we quantified the number of tartrate-resistant acid phosphatase staining+ (TRAP+) osteoclasts and Osteocalcin (OCN)+ osteoblasts (Fig. S7j). TRAP and OCN immunofluorescence staining revealed that Ncstn cKO bones contained significantly more osteoclasts and osteoblasts than controls (Fig. S7k), ruling out a reduction of bone resorption as a cause for the observed phenotype.
Fig. 4
Loss of Notch signaling causes an age and sex-dependent increase in osteogenesis. a 3D-rendered coronal microCT cross-section of Ncstnfl/fl (control) and LeprCre; Ncstnfl/fl (Ncstn cKO) middle-aged mouse femurs. Dotted red box shows isolated metaphyseal trabecular bone. b Quantification of trabecular bone parameters throughout the marrow cavity. BV/TV bone volume/tissue volume, BMD Bone Mineral Density, Tb.Th trabecular thickness, Tb.N trabecular number, Tb.Sp trabecular spacing. Control (n = 9), Ncstn cKO (n = 8). c Coronal microCT images of the femur separated by sex. Blue = male, yellow = female. d Quantification of trabecular bone parameters throughout the marrow cavity separated by sex. Control male (n = 6), control female (n = 3), Ncstn cKO male (n = 6), Ncstn cKO female (n = 5). e Representative immunofluorescent images from the femoral metaphysis of LeprCre; TdTomato and Ncstn cKO ; TdTomato male and female mice showing the contribution of the Lepr lineage to trabecular bone. Trabecular bone is outlined in white and TdTomato+ OCN+ osteoblast lining the bone and TdTomato+ osteocytes which have a spindle appearance within the trabecular bone. Scale bar = 100 um. f Mineral apposition rate (MAR) for control and cKO middle-aged mice with representative imaged, scale bar = 30 um. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.000 1. Data were represented as mean ± s.e.m
Moreover, since hematopoietic cells are produced within the bone marrow, we investigated whether the increase in trabecular bone throughout the marrow cavity, affected the systemic hematopoietic lineage output. Despite the decreased bone marrow cellularity in Ncstn cKO mice (Fig. S7l), the peripheral blood counts did not exhibit any significant change in lineage output (Fig. S7m, n).
SSPCs give rise to both osteoblasts and bone marrow adipocytes.15,35 Typically, osteogenic and adipogenic differentiation potential is inversely correlated.21,51 Consistent with this, in addition to increased bone mineral density, Ncstn cKO mice had fewer adipocytes than controls in all anatomic compartments (epiphysis, metaphysis, diaphysis, periosteum) (Fig. 5a–d). In summary, loss of Notch signaling in SSPCs results in an increase in trabecular bone volume and a reduction in bone marrow adiposity in middle-aged mice and thus essentially prevents age-related skeletal degeneration.
Fig. 5
Loss of Notch signaling causes an age and sex-dependent decrease in adipogenesis. a Histological sections of control and Ncstn cKO middle-aged femurs stained with Movat’s Pentachrome (yellow, bone; red, marrow; blue/green, cartilage). Adipocytes can be seen in the metaphysis as white marrow “ghosts”. Males = blue, females = yellow. b Representative immunofluorescent sections of male and female control and Ncstn cKO middle-aged femurs stained for perilipin (PLIN)+ adipocytes and endomucin (EMNC)+ vasculature. Yellow arrowheads point to characteristic adipocyte accumulation in the metaphysis with aging. SOC secondary ossification center, BM bone marrow, MP metaphysis. Top row, scale bar = 1 000 um. Bottom 2 rows, scale bar = 500 um. c Quantification of PLIN+ adipocytes throughout different compartments of the bone marrow. BM bone marrow, CB cortical bone, PS periosteum. Control male (n = 3), control female (n = 3), Ncstn cKO male (n = 3), Ncstn cKO female (n = 3). d Representative immunofluorescent images from the metaphysis of LeprCre; TdTomato and Ncstn cKO; TdTomato mice. Lepr lineage cells in TdTomato can been seen to give rise to PLIN+ adipocytes by overlap of TdTomato and PLIN (green). Scale bar = 100 um. *P < 0.05, **P < 0.01, ***P < 0.001. Data were represented as mean ± s.e.m
Notch signaling regulates osteo-adipo cell fate decisions in vivoSo far, we showed that Notch signaling in SSPCs controls the homeostatic balance of bone and fat within the skeleton, suggesting that it regulates osteogenic and adipogenic differentiation in vivo. To directly evaluate this, we generated chemical and mechanical skeletal injuries that stimulate adipogenesis and osteogenesis, respectively. To study the adipogenic injury response, we injected Ncstn cKO and control mice with 5-Fluorouracil (5-FU), a drug that is commonly used to treat cancer, that triggers fatty degeneration of the bone marrow similar to what is observed during skeletal aging. Control mice treated with 5-FU exhibited adipocytes distributed throughout the bone marrow (Fig. 6a). However, Ncstn cKO mice showed a significant reduction in bone marrow adiposity compared to controls (Fig. 6a–c). To assess osteogenic regenerative potential, we created mono-cortical tibial defects52,53 in middle-aged mice, which stimulate intramembranous bone formation. At 10 days post-injury, Ncstn cKO mice had a higher bone volume/tissue volume (BV/TV) than control mice, indicating enhanced regeneration (Fig. 6d, e). Thus, loss of Notch signaling suppresses adipogenesis and promotes osteogenesis in vivo in both homeostasis (Figs. 4 and 5) and injury (Fig. 6). Moreover, loss of Notch in SSPCs enhances bone repair in elderly individuals.
Fig. 6
During physical and chemical stress loss of Notch signaling causes decreased adipogenic and increased osteogenic responses. a Representative immunofluorescent images from control and Ncstn cKO mouse femurs 10 days after 5-FU chemical stress known to induce adipogenesis (PLIN+ adipocytes). Scale bar = 1 000 um. b Quantification of PLIN+ staining measured in pixels in the metaphyseal region over the total pixel volume between control (n = 8) and Ncstn cKO (n = 8) mice, (female, yellow; male, blue). c Representative immunofluorescent images from LeprCre; TdTomato and Ncstn cKO; TdTomato mouse femurs after 5-FU treatment showing Lepr lineage cells marked by TdTomato expression giving rise to PLIN+ adipocytes. Scale bar = 1 000 um. d Mono-cortical defect injury microCT 3D reconstruction for middle-aged control and Ncstn cKO mice, post-operative day (POD) 10. e Quantification of trabecular bone parameters for the new woven bone at the injury site formed at POD 10 between control (n = 9) and Ncstn cKO (n = 12) mice. (female, yellow; male, blue). BV/TV bone volume/tissue volume, Tb.Th trabecular thickness, Tb.N trabecular number, Tb.Sp trabecular spacing. *P < 0.05, **P < 0.01, ***P < 0.001. Data were represented as mean ± s.e.m
Ebf3 is a downstream target of Notch signaling in SSPCsWe found that modulating Notch signaling in SSPCs prevented or delayed age-related skeletal degeneration. However, the potential of Notch signaling as a therapeutic target is limited by its known association with cancer54 and the widespread expression of pathway components in different cell types within the skeleton (Fig. S8a) increasing the likelihood of off-target effects. Therefore, we aimed to uncover novel effectors downstream of Notch in SSPCs with a restricted expression pattern. To this end, we isolated LEPR+CD45−CD31−TER-119– SSPCs from middle-aged control and Ncstn cKO mice and performed bulk RNA sequencing to comprehensively analyze the transcriptional changes in SSPCs. We identified 319 genes that were significantly upregulated and 900 that were downregulated in Ncstn cKO SSPCs compared to control SSPCs (P < 0.05) (Fig. S8b). Of these, only 28 were transcription factors and hence have the capacity to control the gene regulatory networks that drive cell fate decisions. We analyzed the expression pattern of each transcription factor in our scRNAseq dataset and pinpointed Early B-cell Factor 3 (Ebf3) as a promising target due to its relatively SSPC-specific expression (Fig. 7a).55Ebf3 was downregulated in Notch signaling-deficient SSPCs (Fig.
Comments (0)