
Remember me
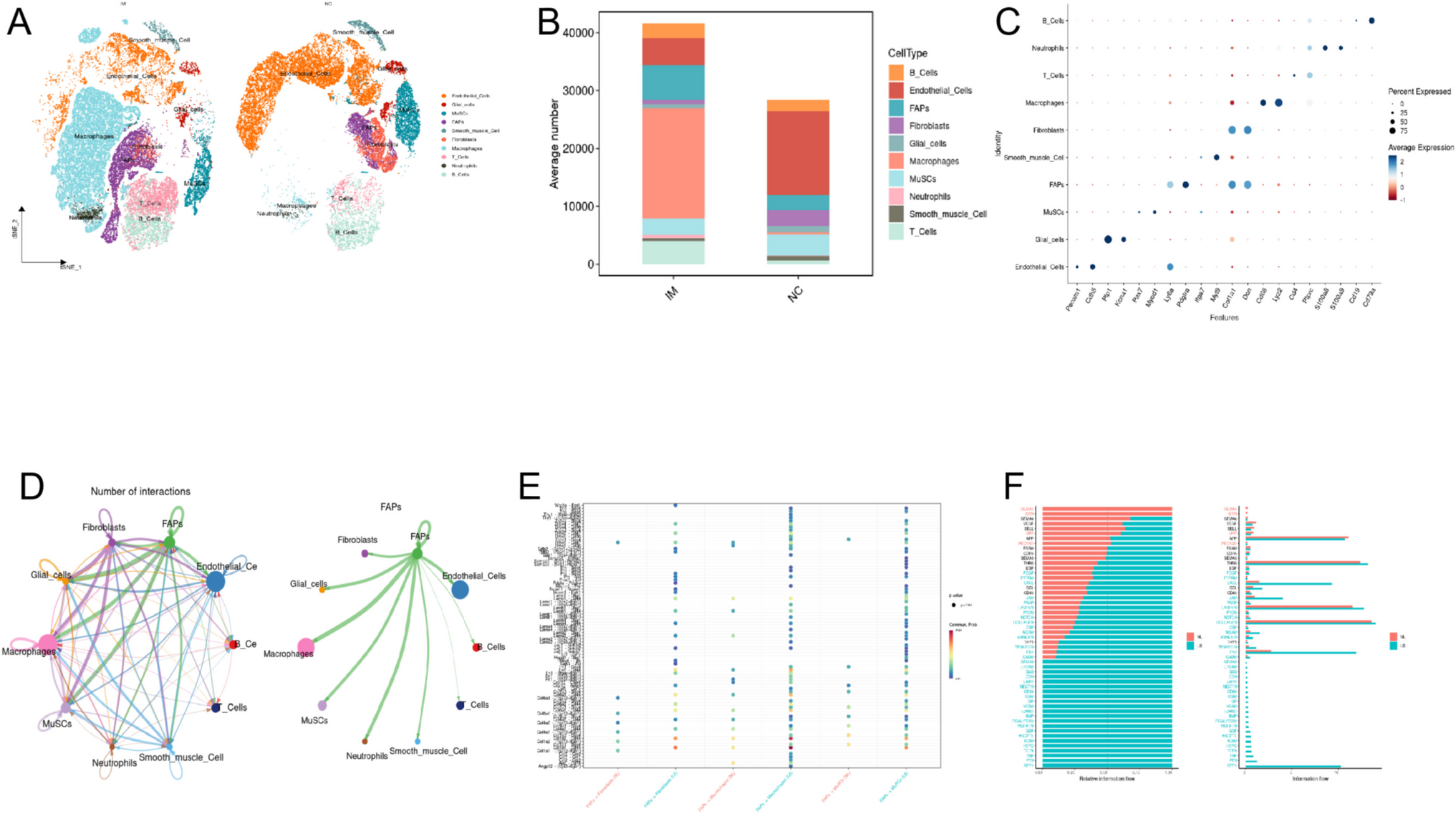
A frequently used model for remyelination research is the dietary cuprizone model (Fig. 1a). The copper chelator cuprizone induces loss of mature OLG, leading to global and maximum demyelination in the CC at 5 weeks cuprizone intoxication. In the CC, OPC activation and recruitment, important for the onset of the remyelination process, typically begins between the third and fifth week of cuprizone feeding. Mature OLG and remyelinated axons are observed within 1 week, and robust remyelination is achieved within 2 weeks after cuprizone withdrawal [22, 24,25,26]. To determine whether TNFR2 stimulation impacts remyelination, saline (PBS) or the TNFR2 agonist EHD2-scTNFR2 were delivered into the brain following 5-week cuprizone-induced demyelination by a single i.c.v. injection (Fig. 1a). Treating early would primarily address effects on demyelination. Since EHD2-scTNFR2 is a human TNFR2 agonist, we used chimeric hu/m TNFR2 knock-in mice, in which the extracellular TNFR2 domain is human and the intracellular domain mouse, maintaining normal mouse TNF signaling and ensuring model suitability [21].
Fig. 1
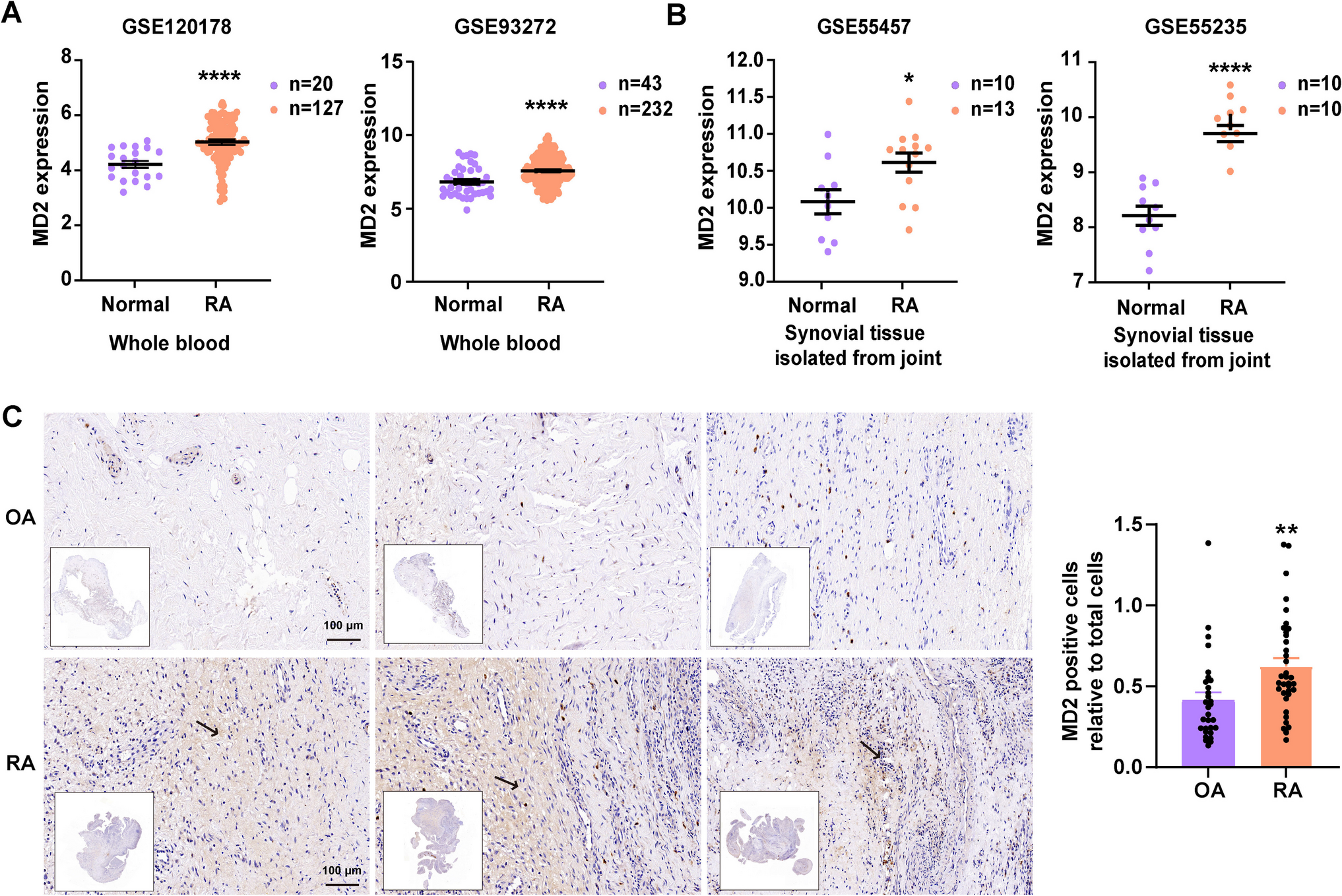
Single EHD2-scTNFR2 treatment following cuprizone-induced demyelination increases myelin density in the corpus callosum over time. a Schematic representation of the experimental design and analyses. To induce demyelination, hu/m TNFR2 knock-in mice were fed for 5 weeks with 0.2% cuprizone. At 5 weeks of cuprizone feeding, mice were injected intracerebroventricularly (− 0.05 anteroposterior: AP; − 0.1 medial lateral: ML; − 0.25 dorsal ventral: DV) with saline (PBS, grey) or the human TNFR2 agonist EHD2-scTNFR2 (R2, blue), fed with normal chow and analyzed 7 or 14 days (DPT) post treatment (DPT). To visualize myelin, coronal brain sections (Bregma 1.10 mm to 0 mm) were stained with Black Gold II (BGII, b) and for myelin basic protein (MBP, d). The optical density of BGII signal (c) and the percentage of MBP coverage (e) in the corpus callosum (CC) were measured. n = 3–4 animals/group, An unpaired t-test (c) or Mann–Whitney U test (e) were used to compare between two treatment groups at the same time point (c, e, not significant), or two time points of the same treatment (c, e, #p < 0.05). Scale bars are 100 µm. a was created using vectors from scidraw.io
To evaluate differences between treatments and the impact of the treatment over time, we examined effects after 1 week (7 days post treatment—DPT) and 2 weeks (14 DPT) after cuprizone removal. To assess the extent of myelin regeneration, the density of myelin in the CC was visualized with a gold-phosphate complex called Black Gold II (BGII) and immunohistochemistry for a major myelin protein, MBP. The optical density of BGII (Fig. 1b,c) and the percentage of MBP coverage (Fig. 1d,e) were slightly, but not significantly, increased at 14 days compared to 7 days post saline treatment, implying ongoing regeneration of myelin between 7 and 14 DPT. Similarly, myelin density increased between 7 and 14 DPT upon EHD2-scTNFR2 treatment, which was significant for MBP coverage (Fig. 1c, e). At both 7 and 14 DPT, the myelin density upon EHD2-scTNFR2 treatment was approximately 10–20% higher than upon PBS treatment, although the difference at each time point did not reach significance (Fig. 1c, e).
To more accurately assess remyelination, ultrastructural parameters of individual (myelinated) axons in large-scale STEM images were analyzed next (Fig. 2a, Table S3). To compare the level of remyelination, we first counted the number of denuded and myelinated axons. At 7 DPT, the percentage of unmyelinated axons was lower in EHD2-scTNFR2-treated animals compared to saline treatment, and similar to 14 DPT (Fig. 2b). At 14 DPT, both upon saline and EHD2-scTNFR2 treatment, approximately 5% of the axons were unmyelinated. Next, we measured the axon diameter of myelinated axons and observed that, at 7 DPT, the average axon diameter was about 70 nm smaller in EHD2-scTNFR2-treated animals without reaching significance, while, at 14 DPT, axon diameter was similar between treatments (Fig. 2c). In comparison with saline, EHD2-scTNFR2 treatment shifted the axon size distribution towards smaller caliber axons at 7 DPT, but not at 14 DPT (Fig. 2d). Furthermore, the myelinated axon area was less following EHD2-scTNFR2 treatment, especially at 7 DPT (Fig. 2e, p = 0.052), while the number of myelinated axons was comparable (Fig. 2f). As changes in axonal size may affect mitochondrial distribution and area [27], we analyzed the number and area of mitochondria in myelinated axons at 7 DPT. In the cross-sectional images, mitochondria in myelinated axons tended to have a larger area upon saline treatment than upon EHD2-scTNFR2 treatment. Conversely, there were no changes in mitochondrial content (% axon area), number of mitochondria per myelinated axon, or percentage of axons with mitochondria between treatments at 7 DPT (Fig. S3a-c). These findings suggest that compared to saline treatment, EHD2-scTNFR2 treatment accelerated the reduction in unmyelinated axons compared to saline treatment, while during early remyelination myelinated axons appear transiently smaller.
Fig. 2
Single EHD2-scTNFR2 treatment following cuprizone-induced demyelination leads to a lower percentage of unmyelinated axons at 7 days post treatment. a Representative STEM images of the corpus callosum of mice treated either with saline (PBS, grey) or EHD2-scTNFR2 (R2, blue) 7 or 14 days (DPT) post treatment (DPT). STEM images are available at full resolution and scale at nanotomy.org. The percentage of unmyelinated axons of total axons (b), axon diameter (c), and myelin thickness were manually measured. The frequency of myelinated axons with different diameters (d) and average myelinated axon area per µm2 (e) were calculated, and myelinated axons per mm2 (f) counted. At least 100 axons per animal were manually measured. n = 3–4 animals/group. An unpaired t-test was conducted to compare between two treatments at the same time point (b, *p < 0.05) or between two time points of the same treatment (b, #p < 0.05). Scale bars are 1 µm
Single TNFR2 agonist treatment following cuprizone-induced demyelination results in thinner myelin sheathsA widely used index to assess myelin is the g-ratio, defined by the ratio between axon diameter and the myelinated fiber diameter (Fig. 3a, b, Table S3). A hallmark of remyelination is the presence of thinner myelin sheaths compared to myelin sheaths generated during developmental myelination [28, 29], resulting in a g-ratio above 0.8 [30]. At 7 and 14 DPT, both small and large caliber axons displayed an average g-ratio around and above 0.8, pointing to remyelinated axons [30].
Fig. 3
Single EHD2-scTNFR2 treatment following cuprizone-induced demyelination results in thinner myelin sheaths. STEM image analysis of the corpus callosum of mice treated either with saline (PBS, grey) or EHD2-scTNFR2 (R2, blue) 7 or 14 days (DPT) post treatment (DPT). From axon and fiber diameters (a), the g-ratio for each axon was calculated, and the g-ratio as average (b) and in relation to average axon diameter are plotted (c). For each animal, the individual myelinated axon diameter was plotted against its myelin thickness (d), highlighting changes in axon size and myelination over time. Solid lines represent the linear (d) regression line. Myelinated axons were scored on their degree of myelin compaction (e, red arrowheads point at non-compacted myelin) and plotted as percentage of the axons analyzed (f). At least 100 axons per animal were manually measured. n = 3–4 animals/group. An unpaired t-test was used to compare between two treatments at the same time point (c, d, *p < 0.05, ****p < 0.00005) or between two time points of the same treatment (c, d, #p < 0.05, ##p < 0.01, ####p < 0.00001). Pearson correlation was used to test the relationship between myelin thickness and diameter of each axon (d). Differences in the degree of myelin compaction were tested with a general linear multivariate model (f, not significant)
At 14 DPT, but not 7 DPT, myelinated axons in EHD2-scTNFR2-treated animals tended to have a higher g-ratio than myelinated axons in saline-treated animals, which reached significance for large caliber axons (between 1 and 2 µm). Interestingly, in the saline-treated group, the g-ratio of large-caliber, and to a lesser extent, small-caliber, axons decreased over time, whereas this was not observed in the EHD2-scTNFR2 group (Fig. 3c), potentially reflecting differences in ongoing myelin membrane wrapping. Furthermore, myelin thickness and axon diameter of each animal correlated for both saline and EHD2-scTNFR2 treatment at 7 and 14 DPT. Correlation coefficients differed significantly between the two treatment groups at 14 DPT and between two time points of the same treatment (Fig. 3d). This further highlights that myelin of particularly larger axons at 14 DPT was thinner upon EHD2-scTNFR2 treatment.
To analyze a higher number of axons, we developed an algorithm which quantifies in STEM images the area covered by myelin and the axon size in an aggregated manner, thus without measuring the properties of each individual axon (Fig. S4a). In line with the manual analysis of individual myelinated axons, at 14 DPT, EHD2-scTNFR2-treated animals tended to have a higher aggregated g-ratio than saline-treated animals (Fig. S4b). Notably, the aggregated g-ratio and axon area values were lower than the manual analysis (Fig. S4b vs Fig. 3b and Fig. S4c vs Fig. 2e), and the aggregated axon area of myelinated axons did not differ between treatments and time points upon EHD2-scTNFR2 treatment (Fig. S4c). The percentage of myelinated area, referring to the area covered by myelin, increased between 7 and 14 DPT in saline-treated animals, whereas it decreased upon EHD2-scTNFR2 treatment (Fig. S4 d).
Thus, both manual and automated analyses signified that axons in EHD2-scTNFR2-treated animals have thinner myelin at 14 DPT. To assess whether myelin was thinner because of a difference in (de)compaction, we next assessed the degree of myelin compaction following a previously described scoring method (Fig. 3e) [31]. Both treatment groups showed similar levels of myelin compaction at 14 DPT (Fig. 3f). However, at 7 DPT, saline-treated animals had around 20% of their axons marginally compacted, while only around 7% of axons of animals treated with EHD2-scTNFR2 have marginally compacted myelin (Fig. 3f). Axons with fully and partially compacted myelin were more abundant in the EHD2-scTNFR2-treated animals at 7 DPT (Fig. 3f), pointing to more myelinated axons being in a later phase of the remyelination process. To assess whether the thinner myelin sheaths in EHD2-scTNFR2-treated animals at 14 DPT resulted from greater compaction, we acquired high-resolution STEM cross-sectional images of large axons at 14 DPT. While large myelinated axons in the EHD2-scTNFR2-treated group may have fewer myelin membrane layers, the myelin sheath compaction appeared more advanced, with clear inner tongues and a more distinct and abundant radial component (Fig. S5) [32]. Therefore, a single i.c.v. injection of a TNFR2 agonist following cuprizone-induced demyelination transiently shifted the myelinated axon size distribution towards smaller caliber axons at early stages of remyelination, which culminated in thinner regenerated myelin sheaths around large-caliber axons at a later stage.
Single TNFR2 agonist treatment following cuprizone-induced demyelination does not induce major changes in microglia and astrocyte abundance in the corpus callosumActivation of microglia and astrocytes is a well-characterized hallmark of cuprizone-induced demyelination [24, 33, 34]. Microglial and astrocyte activation starts as early as 2 weeks of cuprizone feeding, and their activation sustains during remyelination [24]. Therefore, as activated astrocytes and microglia play a crucial role in facilitating remyelination [24] and express TNFR2 in the demyelinated CC [35, 36], we next assessed if the TNFR2 agonist affected the sustained neuroinflammatory response during the remyelination process following cuprizone-induced demyelination. We focused our analysis on the most affected area, the CC, and the whole forebrain, including cortical [37] and basal ganglia [38] regions, relevant areas for neuroinflammatory responses in the cuprizone model. In the CC, the coverage of IBA1-positive microglia notably, but not significantly, decreased between 7 and 14 DPT for both treatments (Fig. 4a, b). Compared to saline, EHD2-scTNFR2 treatment tended to reduce IBA1-positive microglia coverage at 14 DPT in the forebrain (Fig. 4a, c, p = 0.09) and led to a slight, but not significant, decrease in the number of IBA1-positive cells at 14 DPT in the cortex (Fig. 4a, d). Next, we quantified the mRNA expression of pro-inflammatory and anti-inflammatory genes in primary hu/m TNFR2-ki microglia treated with EHD2-scTNFR2 or PBS for 24 h (Fig. S6a). Expression of anti-inflammatory genes such as Arg1 and Il10 was upregulated upon TNFR2 agonist treatment compared to vehicle-treated microglia (Fig. S6c). On the other hand, EHD2-scTNFR2 treatment in vitro did not lead to changes in the expression of pro-inflammatory genes such as Tnf, Il1b, and Nos2 (Fig. S6b).
Fig. 4
Single EHD2-scTNFR2 treatment following cuprizone-induced demyelination does not induce major changes in microglia and astrocyte abundance in the corpus callosum. Representative images of IBA1 (a, microglia marker) and GFAP staining (e, astrocyte marker) of the corpus callosum (CC, red dashed line) and whole forebrain of mice treated with saline (PBS, grey) or EHD2-scTNFR2 (R2, blue) and analyzed 7 or 14 days (DPT) post treatment (DPT). Coronal brain sections were analyzed for the coverage of IBA1 (b, c) or GFAP (f, g), while positive cells were counted in the cortex (CX; d, h). n = 3-4 animals/group. An unpaired t-test (c) or Mann-Witney U test (e) were used to compare between two treatments at the same time point (b–d, f–h, not significant) or between two time points of the same treatment (b–d, f–h, #p < 0.05). Scale bars are 100 µm (overview) and 10 µm (insets)
The distribution of GFAP-positive astrocyte in the CC appeared unaffected (Fig. 4e, f), while the GFAP-positive astrocyte coverage in the forebrain decreased between 7 and 14 DPT (Fig. 4e, g). Similarly, the number of GFAP-positive cells in the cortex decreased for both saline and EHD2-scTNFR2 treatment between 7 and 14 DPT (Fig. 4e, h), indicating that the coverage is likely a reflection of the astrocyte number and not the number and length of their extensions. At both 7 and 14 DPT, no statistically significant difference was observed between treatments. Taken together, TNFR2 agonist treatment does not induce major changes in microglia and astrocyte abundance in the CC.
Single TNFR2 agonist treatment following cuprizone-induced demyelination maintains oligodendrocyte lineage cell numbersRemyelination following demyelination requires OPC recruitment, including proliferation and migration, and their differentiation into myelinating OLG. Within the cuprizone-mediated demyelinated CC, OPC express high levels of TNFR2 [35, 39]. Therefore, we next examined whether EHD2-scTNFR2 modulates remyelination by enhancing OPC recruitment to the demyelinated areas.
While not different at 7 DPT, analysis at 14 DPT revealed that the number of Olig2-positive cells, representing OPC and OLG, was more than threefold higher upon EHD2-scTNFR2 compared to saline treatment both in the mid CC and CC horns (Fig. 5a, b). In fact, while the number of Olig2-positive cells significantly decreased between 7 and 14 days post saline treatment, oligodendrocyte lineage numbers in EHD2-scTNFR2-treated animals were comparable between 7 and 14 DPT, and at 14 DPT significantly higher than in saline-treated animals (Fig. 5b). Co-staining with the proliferation marker Ki67 revealed that the proportion of Olig2-positive cells co-expressing Ki67 was generally low in the CC at 7 and 14 DPT, ranging between 0.5 and 3%, and did not differ between treatments at both time points (Fig. 5c). To assess whether the enhanced numbers of Olig2-positive cells were indeed not a reflection of increased proliferation, we measured the effect of EHD2-scTNFR2 on OPC proliferation in vitro. Naïve primary OPC isolated from hu/m TNFR2-ki mice (P1–P3) treated for 24 h one day after plating or for 48 h immediately after plating with EHD2-scTNFR2 (Fig. S6a) showed no changes in the number of proliferating Olig2-positive cells (Fig. S7a-c). Hence, these findings demonstrate that the number of oligodendrocyte lineage cells in the CC decreased over time for saline treatment but remained high upon single EHD2-scTNFR2 i.c.v. injection without affecting proliferation.
Fig. 5
Single EHD2-scTNFR2 treatment following cuprizone-induced demyelination results in a higher number of oligodendrocyte lineage cells 14 days after treatment. a Representative images of oligodendrocyte lineage marker Olig2 (green) and proliferation marker Ki67 (red) double staining of the corpus callosum (CC) of mice treated either with saline (PBS, grey) or EHD2-scTNFR2 (R2, blue) and analyzed 7 or 14 days (DPT) post treatment (DPT). The orange arrow and rectangle point to a Olig2-positive cells that co-express Ki67, the red arrow and rectangle indicate Ki67-positive cells that are negative for Olig2. The percentage of Olig2-positive cells of total cells in the midCC, CC horns, and total CC (b) and the percentage of Ki67-positive cells of Olig2-positive cells were determined in the total CC (c). n = 3-4 animals/group. An unpaired t-test (c) or Mann-Witney U test (e) were used to compare between two treatments at the same time point (b, c, *p < 0.05, **p < 0.01) or between two time points of the same treatment (b, c, #p < 0.05). Scale bars are 100 µm (overview) and 10 µm (insets)
Comments (0)