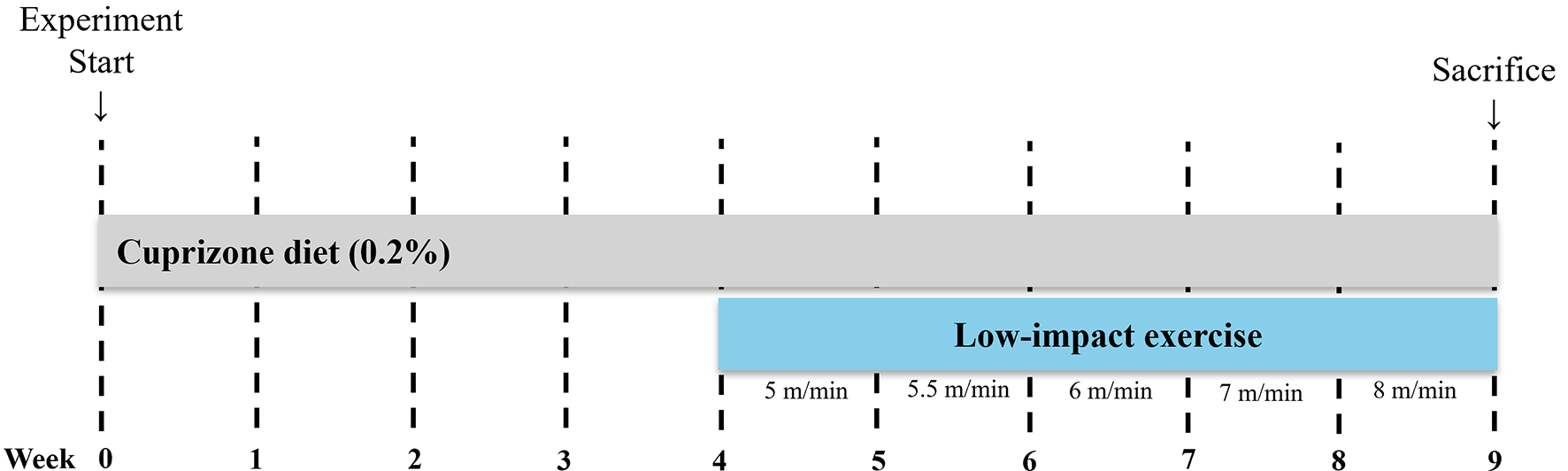
Numerous studies have investigated the effects of physical exercise on neurogenesis in the adult brain [24,25,26]. Exercise is known to stimulate the formation of new neurons in multiple brain regions [27]. For example, Inoue et al. (2015) demonstrated that low-intensity exercise (15 m/min, below the lactate threshold) more effectively promoted adult hippocampal neurogenesis compared to high-intensity exercise (40 m/min, above the lactate threshold) [28]. Similarly, findings from Aguiar et al. suggest that even low-intensity exercise enhances spatial learning and memory [29]. In this study, we investigated whether low-intensity physical exercise could mitigate cuprizone-induced demyelination and neuroinflammation. By employing a low-intensity forced wheel-running model in mice, we sought to determine whether such exercise could counteract the detrimental effects of cuprizone exposure.
Rodent studies indicate that forced exercise, such as treadmill running or wheel running, induces neuroplastic changes in the hippocampus [30], including increased adult neurogenesis [31], enhanced dendritic arborization [32], and greater synaptic plasticity [33]. On the other hand, voluntary wheel running offers advantages such as alignment with innate animal behavior, reduced stress conditions, and minimal researcher interference, thereby leading to robust physiological adaptations such as enhanced endurance and prevention of sarcopenia [34].
Our study confirmed that cuprizone administration significantly decreased body weight, which aligns with findings from previous research [35, 36]. However, low-intensity physical exercise did not substantially influence this cuprizone-induced weight loss. Although exercise provides various health benefits, our data suggest that the observed body weight reduction was primarily due to the systemic effects of cuprizone rather than the exercise intervention itself.
Immunohistochemical examination of hippocampal tissues revealed patterns consistent with previous reports [20]. Specifically, cuprizone-treated mice showed significantly reduced expression of MBP, Ki67, and DCX in the hippocampus compared to sedentary controls [20]. Moreover, cuprizone administration resulted in morphological activation of microglia, as indicated by the increased Iba1 immunoreactivity characterized by hypertrophied cytoplasm and thickened cellular processes. This activation pattern likely reflects the neuroinflammatory responses associated with cuprizone-induced damage [37].
Consistent with other research indicating the benefits of exercise on remyelination [16], our findings demonstrate that low-intensity forced wheel exercise substantially increased MBP expression in the hippocampal regions, including the alveus, SLM, and corpus callosum. This suggests that low-intensity exercise promotes remyelination following cuprizone-induced damage. These observations align with previous findings from Cheng et al., who reported that forced treadmill exercise augmented MBP expression in the penumbra and mitigated demyelination following focal cerebral injury [38]. Additional studies have also reported increased myelin-related protein expression following treadmill exercise, further supporting the beneficial effects of exercise on myelination [39, 40].Nevertheless, these previous studies utilized high-intensity exercise regimens, whereas our study uniquely demonstrated similar neuroprotective effects using a low-intensity exercise paradigm.
In the current study, we observed significant reductions in microglial activation, suggesting that low-intensity exercise exerts anti-inflammatory effects within the hippocampus. This aligns with previous research by Nakanishi et al., who reported that low-intensity exercise suppressed the pro-inflammatory M1 phenotype of microglia and decreased Alzheimer’s disease pathology, including amyloid-beta deposition and neuronal loss [41]. Our findings are also consistent with other studies demonstrating the beneficial effects of exercise on microglial activation and neuroinflammation [42, 43]. To ensure the reliability of our observations, extensive control measures were implemented, including habituation to handling to minimize stress, maintaining consistent dietary conditions, and providing a stable environment.
Prior research has also highlighted potential mechanisms underlying exercise-induced modulation of microglial morphology, such as reduced inflammatory signaling and enhanced neurogenesis [44]. As TNF-α is a key pro-inflammatory cytokine produced by activated microglia and is implicated in the pathogenesis of various neurodegenerative diseases, including MS [45, 46]. In this study, we investigated TNF-α expression in microglia across hippocampal subregions using double immunofluorescence staining with Iba1 and TNF-α.
Our results demonstrate a significant CPZ-induced increase in TNF-α expression and its colocalization with Iba1⁺ microglia, indicating widespread microglial activation. This effect was most prominent in the CA1 region, consistent with regional vulnerability to inflammation. The associated morphological changes, such as hypertrophic and amoeboid shapes, further support the activated state of microglia in the CPZ group. Although TNF-α levels remained elevated in the CPZ + EX group compared to controls, they were notably reduced relative to the CPZ group, particularly in the CA1 region where a significant decrease in TNF-α/Iba1 colocalization was observed. These findings suggest that voluntary exercise may attenuate microglial inflammation, in part, by modulating TNF-α expression.
Furthermore, colocalization analysis revealed significantly higher TNF-α expression in Iba1-positive cells in both CPZ and CPZ + EX groups compared to controls, highlighting the continued inflammatory potential of microglia even following exercise intervention. This differential regulation suggests that exercise may selectively influence certain aspects of microglial activation while leaving others, such as cytokine production, relatively unaffected.
While activated microglia represent a major source of TNF-α the persistence of TNF-α despite reduced Iba1 expression implies contributions from other CNS cells such as astrocytes or infiltrating immune cells [47]. Temporal dynamics may also account for these findings; previous studies indicate that morphological changes in microglia may precede reductions in cytokine production [48]. Additionally, exercise may induce a shift toward an intermediate microglial state that retains cytokine expression while modifying other functional properties, as observed in Alzheimer’s models [49].
This dissociation between morphology and cytokine levels parallels findings by Xu et al., who reported that specific signaling pathways independently regulate microglial morphology and inflammatory output [50]. The high colocalization of TNF-α with Iba1 in both CPZ and CPZ + EX groups suggests that while microglial density may decline, remaining activated microglia maintain robust TNF-α expression. These findings underscore the complexity of microglial responses and suggest that broad suppression of microglial activation may not suffice to reduce pro-inflammatory cytokine levels. Instead, targeted interventions focusing on cytokine-specific regulatory mechanisms—such as the NLRP3 inflammasome or MAPK pathways—may offer greater therapeutic efficacy [51, 52]. Future studies should investigate whether alternative exercise regimens or combined therapeutic approaches could better modulate inflammatory responses in demyelinating disorders.
Recent evidence underscores the significant role of microglial innate immune memory in influencing neurological disease pathology. As the resident immune cells of the central nervous system (CNS), microglia can acquire a memory-like state in response to peripheral inflammatory stimuli such as lipopolysaccharide exposure. This phenomenon, mediated through mechanisms of trained immunity and immune tolerance, involves epigenetic reprogramming that alters microglial function over the long term [53, 54]. Trained immunity may result in an enhanced pro-inflammatory response upon re-exposure to stimuli, while immune tolerance is associated with diminished reactivity [55]. These persistent changes in microglial activation can significantly affect CNS homeostasis and contribute to the progression of neurological disorders [53, 54]. For instance, trained microglia have been shown to exacerbate neurodegeneration in Alzheimer’s disease by facilitating β-amyloid accumulation and sustaining chronic neuroinflammation [55, 56]. In contrast, microglial immune tolerance may confer neuroprotective effects by attenuating inflammatory responses under certain conditions. In the cuprizone model employed in this study, peripheral inflammatory cues could influence microglial activation states, thereby modulating processes such as hippocampal demyelination and adult neurogenesis [55, 57]. Exploring how microglial innate immune memory interacts with exercise-based interventions may offer novel insights into therapeutic strategies for neurological diseases like MS, where the interplay between peripheral immune activation and CNS pathology is of critical importance [53, 58].
Interestingly, our low-intensity exercise intervention did not significantly improve neurogenesis markers, such as DCX and Ki67, which might be attributed to the severity of cuprizone-induced damage. It is possible that the moderate intensity of our exercise regimen was insufficient to stimulate neurogenesis significantly, given that more vigorous physical activity has been associated with more pronounced neuroplastic adaptations [31]. Indeed, while cuprizone administration significantly impaired hippocampal neurogenesis, low-intensity physical exercise did not effectively reverse this effect.
However, in our study, low-intensity physical exercise did not significantly improve markers of neurogenesis such as DCX and Ki67. This discrepancy could be attributed to the severity of cuprizone-induced hippocampal damage, which might have been too extensive to be significantly reversed by the moderate-intensity exercise employed. Indeed, previous research has shown that higher-intensity exercise regimens are typically more effective at eliciting neuroplastic adaptations [59]. While cuprizone administration significantly impaired hippocampal neurogenesis, low-intensity exercise was insufficient to fully counteract this effect in our model.
The co-expression of Ki67 and NeuN in our study indicates that actively dividing cells are differentiating into neurons, a process crucial for maintaining neural plasticity and cognitive function [60]. This finding is particularly relevant in the context of the cuprizone model, where neurogenesis is often compromised due to demyelination and neuroinflammation [61]. Distinguishing newly formed neurons from other dividing cells, such as glial cells, is essential for accurate interpretation of neurogenesis data [62]. The use of NeuN helps confirm the neuronal identity of Ki67-positive cells, ensuring that the observed neurogenesis reflects neuronal differentiation rather than glial cell proliferation. This specificity is crucial for understanding the true impact of our interventions on neuronal populations and their potential to restore or enhance neurogenesis in conditions where it is impaired.
The observed differences in Ki67+/NeuN + cells following exercise suggest that physical activity can induce neuroplastic adaptations by promoting neurogenesis. This modulation could have significant implications for hippocampal function and behavior, given the well-established link between neurogenesis and learning and memory [60]. Further investigation into the mechanisms underlying these effects could provide insights into how to enhance or restore neurogenesis in conditions where it is impaired, such as neurodegenerative diseases or psychiatric disorders.
Our study highlights several limitations that warrant consideration. First, we exclusively used male mice to control variability associated with the estrous cycle and to maintain consistency with prior research. However, sex differences play an important role in neuroinflammation and demyelination processes [34, 63], emphasizing the need for future studies incorporating both male and female subjects to improve generalizability and explore potential sex-specific therapeutic strategies.
Second, we evaluated only one intensity of exercise, limiting our ability to compare the differential effects of various exercise regimens. The expression of brain-derived neurotrophic factor, known to increase myelin component proteins, is influenced by exercise intensity. Although low-intensity exercise has been shown to promote neurogenesis in the hippocampus, its effect may be less pronounced than that of higher-intensity exercise [64]. Future studies should explore a range of exercise intensities to determine the optimal conditions for neuroprotection and remyelination in demyelinating conditions.
Third, our study specifically focused on the hippocampus, while demyelination affects multiple brain regions. Examining other areas, such as the cerebellum, cortex, or spinal cord, could provide a more comprehensive understanding of how exercise influences neuroprotection across the central nervous system.
Additionally, we assessed only short-term exercise effects. Long-term studies are necessary to elucidate the cumulative or delayed benefits of low-intensity exercise on neurogenesis, remyelination, and neuroinflammation in demyelinating conditions.
Finally, we acknowledge the absence of rescue experiments, which could have provided deeper mechanistic insights into functional recovery. Future studies should investigate whether the neuroprotective effects of exercise persist after cessation or if additional interventions, such as pharmacological treatments or neurotrophic factor administration, could enhance recovery. Exploring the optimal duration and intensity of exercise in combination with other therapeutic strategies will be crucial for translating these findings into clinical applications.
In conclusion, our study demonstrates that low-intensity physical exercise enhances remyelination and modulates neuroinflammatory processes in a cuprizone-induced demyelination model. Specifically, low-intensity exercise significantly increased MBP expression and reduced microglial activation, indicating beneficial neuroprotective effects. Although exercise did not significantly impact neurogenesis markers, these findings highlight its potential therapeutic role for managing demyelinating neurological disorders. Future research should further explore the detailed mechanisms underlying these effects and evaluate different intensities and durations of exercise interventions to maximize clinical relevance.

Comments (0)