
Remember me

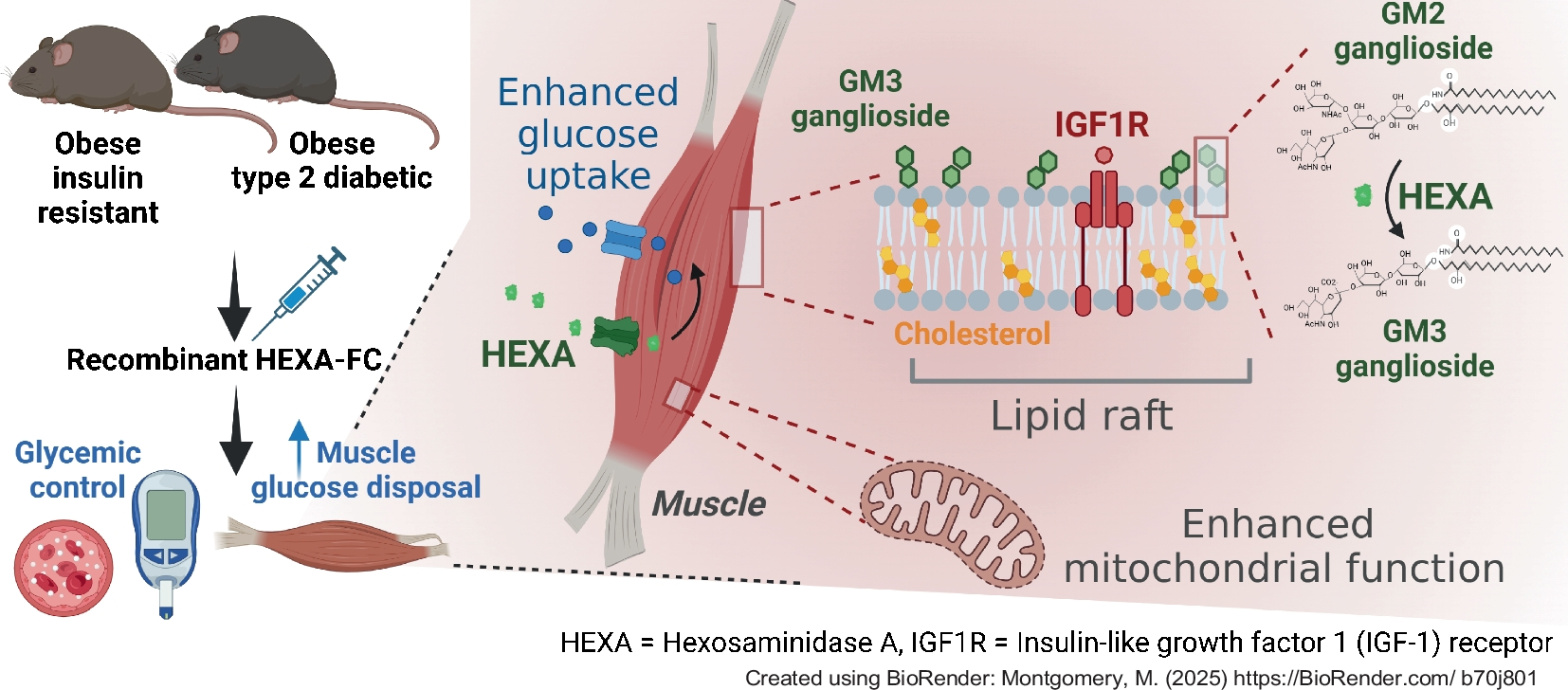

We have recently shown that daily administration of HEXA recombinant protein (1 mg/kg body weight) for 4 weeks in obese mice improves glycaemic control by enhancing glucose disposal into skeletal muscle [4]. While these data are important as proof-of-concept for the therapeutic utility of HEXA, circulating HEXA is rapidly degraded, with plasma levels returning to baseline 4 h following injection [4]. Therefore, we produced a long-acting HEXA-FC fusion protein that is still detected in the circulation 3 days post-injection (Fig. 1a) to allow for more therapeutically relevant dosing.
To evaluate the effects of chronic HEXA-FC on glycaemic control, male C57BL/6 mice were fed a high-fat diet (HFD) for 4 weeks, followed by bi-weekly administration of HEXA-FC or FC (1 mg/kg body weight) for a further 8 weeks. HEXA-FC treatment led to a significant increase in plasma HEXA (Fig. 1b–d), as well as increased HEXA content in quadriceps muscle but not liver (Fig. 1e, f), which is not surprising given that circulating proteins bearing mannose-6-phosphate residues, as is the case for HEXA, can be internalised through the mannose-6-phosphate receptor [23, 24]. Of note, HEXA was not detected in adipose tissue or heart from these mice, which is supported by the finding that HEXA is only expressed in liver and skeletal muscle, but not adipose tissue, heart, kidney, spleen, lung or brain (Fig. 1g).
HEXA-FC treated mice showed a significant reduction in random blood glucose (Fig. 1h), a mild decrease in fasting blood glucose (Fig. 1i) and improved glucose tolerance (Fig. 1j). While fasting plasma insulin was unaffected, there was a significant reduction in glucose-stimulated plasma insulin (Fig. 1k). Insulin tolerance was not impacted by HEXA-FC treatment (Fig. 1l). Together, these data demonstrate that HEXA-FC improves glycaemic control independently of insulin.
Chronic HEXA-FC increases peripheral glucose disposal into skeletal muscleTo understand whether the improvements in glucose tolerance were due to an increase in peripheral glucose disposal (primarily into muscle) or a suppression of endogenous glucose production/output (primarily by the liver), we investigated glucose homeostasis using a stable isotope-labelled GTT. HEXA-FC treatment tended to increase peripheral glucose disposal (p=0.074, Fig. 2a) but did not affect endogenous glucose output (Fig. 2b), indicating a potential impact of HEXA-FC on muscle glucose metabolism. To confirm this, mice underwent an i.p. GTT with simultaneous administration of radiolabelled 2DG (Fig. 2c and electronic supplementary material [ESM] Fig. 1a), which allowed assessment of glucose uptake into tissues. HEXA-FC treatment tended to increase glucose uptake in quadriceps and gastrocnemius muscle (p=0.0616, Fig. 2d), but not in adipose tissue or kidney, while glucose uptake was reduced in the heart (Fig. 2e). The reduction in cardiac glucose uptake was independent of changes in markers of cardiac stress/injury (atrial natriuretic peptide [ANP] and b-type natriuretic peptide [BNP]) and pathological cardiac hypertrophy (shift in α- to β- myosin heavy chain) (ESM Fig. 1b). We further observed enhanced Akt S473 phosphorylation in muscle but not adipose tissue, heart or liver (Fig. 2f–h), as well as increased glucose oxidation in EDL muscle (Fig. 2i) and a non-significant increase in soleus muscle (p=0.092, Fig. 2j). Fatty acid oxidation (Fig. 2k, l) and fatty acid uptake (ESM Fig. 1c, d) in EDL and soleus muscles did not differ between groups.
HEXA-FC treatment did not impact body weight, fat or lean mass (ESM Fig. 1e–g), any tissue weights (ESM Fig. 1h–k) or whole-body oxygen consumption (ESM Fig. 1l), while the respiratory control ratio was reduced (ESM Fig. 1m), suggesting an increase in systemic fat oxidation, potentially due to an increase in hepatic fatty acid oxidation, as recently shown [22]. Food intake was not different between groups but locomotor activity was mildly reduced (ESM Fig. 1n, o). Together, these results show that HEXA-FC improves glycaemic control by enhancing peripheral glucose disposal into skeletal muscle.
HEXA-FC treatment increases muscle GM3 and expression of IGF1 target genesWe next aimed to elucidate the mechanism underlying the metabolic impact of HEXA-FC on skeletal muscle. Targeted LC/MS ganglioside analysis in quadriceps muscle identified 17 GM3 ganglioside species, with an overall non-significant 42% increase in total GM3 content (p=0.067, Fig. 3a) and a significant increase in specific GM3 species following HEXA-FC treatment (Fig. 3b). Untargeted lipidomics analysis identified 1132 lipid species, with no differences between groups (Fig. 3c, d). However, although hexosyl ceramides (direct products of GM3 degradation) were not different (Fig. 3e), we did observe a significant increase in muscle ceramide content (Fig. 3f). This highlights a specific effect of HEXA-FC on sphingolipid remodelling in muscle.
Fig. 3
HEXA-FC treatment remodels the skeletal muscle lipidome and proteome. C57BL/6 mice were fed an HFD for 4 weeks followed by bi-weekly administration of FC or HEXA-FC (1 mg/kg body weight) for a further 8 weeks. (a, b) Total GM3 content (p=0.067) (a) and GM3 lipid species (b) in quadriceps muscle (n=6–7 per group). (c, d) Untargeted lipidomics analysis in quadriceps muscle: (c) principal component analysis and (d) volcano plot analysis, highlighting non-significant changes in ceramide (CER), hexosyl ceramides (HexCer) and triglycerides (TAG) (n=7 per group). (e, f) Total quadriceps (Quad) muscle content of (e) hexosyl ceramides and (f) ceramides (n=7 per group). (g–j) Untargeted proteomics analysis in skeletal muscle: (g) principal component analysis, (h) volcano plot analysis, highlighting mitochondrial proteins, (i) analysis of the major pathways associated with proteins increased in muscle following HEXA-FC treatment, and (j) volcano plot analysis, highlighting IGF1 targets (n=6–7 per group). Values are means ± SEM. *p<0.05 vs FC control, as assessed using two-tailed unpaired t tests (a) or two-way ANOVA and Bonferroni post hoc analysis (b). For the volcano plots in (h) and (j), dark dots indicate significant changes with HEXA-FC treatment and light dots indicate non-significant changes
Untargeted proteomics analysis identified 1611 proteins and revealed distinct changes in protein expression in the muscles of mice treated with FC or HEXA-FC (Fig. 3g). Of those, 288 proteins were significantly increased and 64 proteins were reduced following HEXA-FC treatment (Fig. 3h). Interestingly, of the 288 proteins that were increased, 185 proteins were of mitochondrial origin (64%), while only five mitochondrial proteins were reduced in the muscles of HEXA-FC treated mice (Fig. 3h). Supporting these findings, pathway analysis highlighted activation of pathways associated with ‘aerobic respiration’, ‘respiratory electron transport’ and ‘metabolism of proteins and lipids’ (Fig. 3i). We have previously shown that HEXA exacerbates IGF1-mediated signalling, which was associated with increased glucose disposal into muscle [4]. Here, we recapitulated this finding, showing marked upregulation of 47 IGF1 receptor (IGF1R) target genes [25] after HEXA-FC treatment (Fig. 3j). These data highlight that HEXA-FC exacerbates IGF1 receptor-mediated protein expression, which is likely to mediate the observed enhancement of muscle glucose uptake, as previously shown [4].
Glycaemic improvements with HEXA-FC are recapitulated in a mouse model of type 2 diabetesWe next assessed the efficacy of HEXA-FC in a mouse model of severe obesity and type 2 diabetes. db/db mice received HEXA-FC or FC bi-weekly (1 mg/kg body weight) for a total of 8 weeks. While HEXA-FC treatment did not significantly increase plasma HEXA (Fig. 4a), there was a substantial increase in HEXA within skeletal muscle, heart and liver, but not adipose tissue (Fig. 4b, c). Notably, HEXA was barely detectable in any tissue except adipose tissue in mice receiving the FC control.
Fig. 4
HEXA-FC treatment improves glycaemic control in mice with severe obesity and type 2 diabetes. db/db mice received bi-weekly administration of FC or HEXA-FC (1 mg/kg body weight) for a total of 8 weeks. (a) Plasma HEXA as assessed by ELISA, and (b, c) immunoblotting analysis of HEXA in quadriceps muscle, heart, liver and epididymal adipose tissue of db/db mice (n=4–5 per group). (d) Random blood glucose over the last 5 weeks of treatment, (e) fasting blood glucose, (f) glucose tolerance, (g, h) fasting and glucose-stimulated plasma insulin (HEXA effect in h, p=0.089), and (i) insulin tolerance of FC control and HEXA-FC treated mice (n=6 per group). Values are means ± SEM. *p<0.05 vs FC control, as assessed using two-tailed unpaired t tests (a, c, e, g) or two-way ANOVA and Bonferroni post hoc analysis (d, f, h, i)
HEXA-FC treatment led to a pronounced reduction in random blood glucose from a mean of 41 mmol/l in FC-treated mice to 29 mmol/l in HEXA-FC treated mice (Fig. 4d). Importantly, HEXA-FC blunted the age-related increase in blood glucose in db/db mice (Fig. 4d). HEXA-FC treatment resulted in a 40% decrease in fasting blood glucose (Fig. 4e) and markedly improved glucose tolerance (Fig. 4f). Surprisingly, HEXA-FC increased fasting plasma insulin (Fig. 4g) and tended to increase glucose-stimulated plasma insulin (p=0.089, Fig. 4h), but insulin tolerance did not differ between groups (Fig. 4i). Together, these results demonstrate that HEXA-FC treatment improves glycaemic control in a mouse model of severe type 2 diabetes.
HEXA-FC treatment enhances muscle glucose disposal and increases muscle size in db/db miceUsing a stable isotope-labelled GTT, we found that HEXA-FC exacerbated peripheral glucose disposal (Fig. 5a) but did not affect endogenous glucose output (Fig. 5b). A GTT with radiolabelled 2DG (ESM Fig. 2a) confirmed a pronounced improvement in glucose tolerance (Fig. 5c), with a concomitant increase in glucose uptake into quadriceps and gastrocnemius muscle (Fig. 5d), as well as epididymal and inguinal adipose tissue (Fig. 5e), but not heart or kidney (ESM Fig. 2b). This was accompanied by increased Akt S473 phosphorylation in muscle, adipose tissue and heart, but not liver (Fig. 5f–h), as well as increased glucose oxidation in EDL muscle (Fig. 5i). Furthermore, HEXA-FC mice exhibited increased fatty acid oxidation (Fig. 5j) but not fatty acid uptake (Fig. 5k) in EDL muscle. While body weight (ESM Fig. 2c) and fat mass (ESM Fig. 2d) were not different, HEXA-FC treatment increased lean mass (Fig. 5l) and the size of individual muscles, including the quadriceps and gastrocnemius muscles (Fig. 5m).
Fig. 5
HEXA-FC treatment increases muscle glucose disposal in mice with severe obesity and type 2 diabetes. db/db mice received bi-weekly administration of FC or HEXA-FC (1 mg/kg body weight) for a total of 8 weeks. (a, b) Mice underwent a stable isotope-labelled GTT, followed by assessment of (a) exogenous glucose load and (b) endogenous glucose output (n=4 per group). (c–e) An i.p. GTT was performed with simultaneous administration of radiolabelled 2DG. Mice were killed 30 min after glucose administration and tissues were assessed for 2DG uptake: (c) glucose tolerance, (d) glucose uptake into quadriceps (Quad) and gastrocnemius (Gastroc) muscle, and (e) glucose uptake into epididymal adipose tissue (Ep.AT) and inguinal adipose tissue (Ing.AT) (n=5–6 per group). (f–h) Akt S473 phosphorylation in (f) quadriceps muscle and (g) epididymal adipose tissue, heart and liver; (h) quantification of the immunoblots shown in (f) and (g) (n=4–5 per group; p=0.086 for heart). (i–k) Glucose oxidation (i), fatty acid (FA) oxidation (j) and fatty acid uptake (k) in EDL muscle (n=4–6 per group). (l) Systemic lean mass and (m) individual muscle weights of quadriceps muscle (Quad), gastrocnemius muscle (Gas) and the tibialis anterior (Tib) (n=5–6 per group). Values are means ± SEM. *p<0.05 vs FC control, as assessed using two-tailed unpaired t tests (h–m) or two-way ANOVA and Bonferroni post hoc analysis (a–e)
HEXA-FC treatment increases muscle GM3 content and mitochondrial proteinsHEXA-FC treatment significantly increased total GM3 content (Fig. 6a) and GM3 ganglioside species (Fig. 6b). Proteomics analysis identified 1622 proteins, and revealed distinct HEXA-FC mediated changes in the muscle proteome (Fig. 6c, d), particularly a marked increase in mitochondrial proteins (Fig. 6d), with 58 mitochondrial proteins (62%) being significantly increased in HEXA-FC muscle, including subunits of complex I (25 subunits) of the electron transport chain (Fig. 6d). Reactome pathway analysis highlighted activation of pathways associated with ‘aerobic respiration’, ‘respiratory electron transport’ and ‘complex I biogenesis’ (Fig. 6e). Surprisingly, there were only modest changes in IGF1 receptor target genes (Fig. 6f), which may be related to low IGF1 receptor expression in db/db muscle [26]. However, Qiagen ingenuity pathway analysis revealed that HEXA-FC treatment regulated proteins associated with ‘eukaryotic translation’, ‘eukaryotic initiation factor 2 signalling’ and ‘ribosomal quality control’ (Fig. 6g), pathways that are important for protein synthesis and may contribute to the increase in muscle mass [27]. In addition, upstream regulators predicted to drive these effects included c-myc (MYC), n-myc (MYCN) and the insulin receptor (Fig. 6h), which are known regulators of protein synthesis [28, 29] and could contribute to the increase in muscle mass.
Fig. 6
HEXA-FC treatment remodels the skeletal muscle lipidome and proteome in db/db mice. db/db mice received bi-weekly administration of FC or HEXA-FC (1 mg/kg body weight) for a total of 8 weeks. (a, b) Total GM3 content (a) and GM3 lipid species (b) in quadriceps muscle (n=4 per group). (c, d) Untargeted proteomics analysis in skeletal muscle: (c) principal component analysis, (d) volcano plot analysis, highlighting mitochondrial proteins. (e) Reactome pathway analysis of the major pathways associated with proteins increased in muscle following HEXA-FC treatment. (f–h) volcano plot analysis highlighting IGF1R targets (f), and Qiagen ingenuity pathway analysis of canonical pathways (g) and predicted upstream regulators (h) (n=3 per group). (i–k) Untargeted lipidomics analysis in quadriceps muscle: (i) principal component analysis, (j) volcano plot analysis highlighting changes in ceramide (CER), hexosyl ceramides (HexCer) and triglycerides (TAG); (k) total cardiolipin content in the quadriceps muscle (Quad) (n=4 per group). Values are means ± SEM. *p<0.05 vs FC control, as assessed using two-tailed unpaired t tests (a, k) or two-way ANOVA and Bonferroni post hoc analysis (b). For the volcano plots in (d) and (f), dark dots indicate significant changes with HEXA-FC treatment and light dots indicate non-significant changes. EIF2, eukaryotic initiation factor 2; INSR, insulin receptor; MYC, c-myc; MYCN, n-myc; NFE2L2, nuclear factor erythroid 2-related factor 2; ROBO, roundabout
While lipidomics analysis did not point to any substantial changes in the muscle lipidome following HEXA-FC treatment (Fig. 6i, j), we observed a significant increase in the level of cardiolipin, a phospholipid that is localised to the inner mitochondrial membrane (Fig. 6k), suggesting that HEXA-FC treatment may increase mitochondrial capacity in db/db muscle. In addition, various sphingolipids, including ceramides and sphingosine, were mildly increased in HEXA-FC muscle (Fig. 6j), highlighting mild remodelling of sphingolipid metabolism beyond the expected increases in GM3 gangliosides.
Acute HEXA treatment inhibits insulin secretion and remodels lipid raft composition within pancreatic isletsHEXA-FC reduced plasma insulin in HFD mice (Fig. 1k) but tended to increase plasma insulin in db/db mice (p=0.089, Fig. 4h). To understand whether HEXA has direct effects on insulin secretion, we isolated pancreatic islets from lean C57BL/6 mice and incubated them with HEXA protein. Interestingly, while HEXA inhibited glucose-stimulated insulin secretion after acute treatment (10 ng/ml, 1 h), this effect was lost after chronic 24 h incubation (Fig. 7a). The acute inhibition of insulin secretion was accompanied by an increase in intracellular insulin content (Fig. 7b).
Fig. 7
HEXA suppresses insulin secretion and remodels lipid rafts in pancreatic islets. (a, b) Pancreatic islets were isolated from lean C57BL/6 mice and subjected to acute (1 h) or chronic (24 h) incubation with HEXA protein (10 ng/ml), followed by assessment of glucose-stimulated insulin secretion (a) and intracellular insulin content (b) (n=5 per group). (c–h) Pancreatic islets were isolated from HFD mice chronically treated with HEXA for 4 weeks, followed by assessment of the proteome of whole islets and fractionated lipid rafts: (c) principal component analysis and (d) volcano plot analysis of whole islets, and (e) principal component analysis and (f) volcano plot analysis of fractionated islet lipid rafts; (g) proteins that were uniquely present only in control lipid rafts or in lipid rafts of HEXA-treated mice (PKP1, plakophilin-1; SYP, synaptophysin), and (h) pathways associated with the proteins that were absent in pancreatic islet lipid rafts following HEXA treatment (n=4–5 per group). Values are means ± SEM. A statistically significant HEXA-FC treatment effect (p<0.05) is indicated in (a, b) as assessed by two-way ANOVA and Bonferroni post hoc analysis
Given that HEXA inhibited insulin secretion in isolated islets, and following on from our previous observation that chronic HEXA-FC administration reduced plasma insulin in HFD mice, we next took advantage of isolated pancreatic islets obtained from HFD mice chronically treated with HEXA [4], and assessed the proteome of fractionated lipid rafts from these islets using the method described previously [4, 22]. While the whole-islet proteome (3541 proteins identified, Fig. 7c, d) and lipid raft proteome (150 proteins identified, Fig. 7e, f) were similar between groups, we found that 44 proteins were absent from lipid rafts following HEXA treatment (Fig. 7g), and these proteins were associated with the immune system and cellular stress (Fig. 7h), suggesting that HEXA may suppress stress and the immune response within pancreatic islets.
Comments (0)