We observed a broad metabolomic signature associated with the intervention, with differences observed in metabolites from every one of the biochemical classes (or super pathways) represented. Under a conservative correction for multiple testing, 26% of NMR-derived metabolites and 12% of MS-derived metabolites were altered by the intervention, suggesting that a sizeable proportion of metabolite changes in type 2 diabetes are modifiable. We report a lipid pattern change with reduction in TG-rich lipoproteins across the lipoprotein cascade, but enrichment of (lyso)plasmalogens and reversal of amino acid changes associated with type 2 diabetes, as well as a reduction in a range of sugars beyond glucose, including fructose and mannose.
A major strength of this study is the use of samples and clinical data collected from a relatively large (compared with existing literature) cluster randomised trial with a well-matched control arm. Measuring metabolites both at baseline and at 12 months added to the robustness of the analysis while the use of two complementary metabolomics platforms increased the overall coverage of the metabolome beyond that which has been evaluated previously. However, the MS data are semi-quantitative meaning that these findings require further validation using targeted techniques to allow absolute quantification. Twelve-month samples were not available from participants who dropped out of the trial; this was only a small number and since the primary analyses concerned paired baseline and 12 month measures, bias from this differential missingness was minimised. While our study design enabled us to conduct a thorough evaluation of the metabolomic impact of the Counterweight-Plus intervention overall, it is challenging to attribute those changes to specific elements of the intervention (e.g. to fat loss per se or ‘upstream’ changes in diet). Results from attempts here to extract the weight-loss effects should be interpreted with caution, especially given that these analytical manoeuvres alter the trial structure and, as such, have the potential to introduce bias.
Characteristic changes in metabolite profile
Many of the metabolites influenced by the intervention have also been identified as potential risk factors for type 2 diabetes development with opposite and proportionate effect sizes. For example, we see decreased concentrations of BCAAs following dietary intervention where plasma concentrations of these BCAAs are frequently elevated in type 2 diabetes [5, 9, 21]. The decrease we observe in BCAAs largely agrees with existing findings from smaller, targeted studies of weight-loss interventions [14, 22, 23]. While not all studies report this decrease in BCAAs after diet-induced weight loss [24], this may be due to a lack of power to discern what is likely to be a smaller effect than that from equivalent analyses considering surgical interventions. By using an untargeted metabolome-wide approach, in this work we were able to further characterise the plasticity of this highly relevant network. We observed concurrent decreases in several γ-glutamyl BCAA dipeptides allocated to the same cluster as the BCAAs themselves; γ-glutamyl amino acids are produced when the enzyme, γ-glutamyl transpeptidase, present mainly in the liver, catalyses the transfer of the γ-glutamyl moiety of glutathione to an amino acid [25].
Similarly to BCAAs, the reduction we see in the levels of several simple sugars, including the monosaccharides fructose, glucose and mannose, are opposite to the elevations seen in levels of these metabolites in the presence of obesity. While structurally similar, the predominant dietary sources, metabolic pathways and biological effects of these simple sugars are quite different though interdependent [26]. Mannose specifically has been associated not only with insulin resistance but also with higher risk of several chronic diseases including type 2 diabetes and CVD [27]. The existing literature concerning the most strongly associated metabolite from the MS dataset, erythronate, is limited; however, further insight into its relevance to type 2 diabetes can be gained by considering its relationship with other measured metabolites. In our data, erythronate sits in a cluster with (i.e. is correlated with) several common sugar alcohols (ribitol, erythritol, arabitol/xylitol) that can be found naturally in fruits but that are also commonly used as artificial sweeteners. Erythritol is predominantly excreted in urine, with the remaining 5–10% being oxidised to erythronate [28]. While designated as a xenobiotic in the Metabolon data and previously thought not to be produced endogenously [29], erythritol may in fact be produced endogenously from glucose [30]. This, together with preliminary evidence showing an association between erythritol and adiposity gain in young adults [30], serves to contextualise our findings of reduced levels of erythronate and related metabolites (ribitol, orotidine and erythritol all had p<0.05 before Holm-correction) in participants in the intervention group.
Participants in the intervention arm saw increases in several lipids previously associated with a favourable metabolic profile. Specifically, increases were seen in concentrations of several (lyso)plasmalogens, a special class of phospholipids characterised by the presence of a vinyl–ether bond at the sn-1 position. In a cross-sectional study of participants with overweight and obesity, plasmalogen levels were found to be inversely correlated with body fat percentage but seemingly not related to BMI or WHR [31]. The lack of association for these commonly used indicators of adiposity may be related to their suboptimal performance as proxies for adiposity in this relatively small sample of individuals all with BMI>25 kg/m2 (n=65). Alternatively, this may point towards a more complex interplay between metabolic health and plasmalogens.
Detecting associations with changes in exogenous factors
While changes to the metabolism can be expected in response to the intervention-induced weight loss experienced by many of those in the intervention group, we also expect the adoption of new dietary patterns and a change in medication regimes. At a metabolomic level, and notwithstanding possible limitations linked to limits of detection, we assume that, where we see high levels of missingness for given metabolites, these patterns are indicative of absence and/or very low concentration. This does mean that naive application of a linear model will be underpowered to detect mean differences in concentrations. However, examining between-group differences in presence/absence can allow detection of meaningful relationships. To this end, the logistic regression analysis here revealed between-group differences in the frequency of detection of both potential dietary biomarkers and medications. For example, S-allylcysteine, a proposed biomarker for garlic consumption [32], was enriched in the intervention group while ethyl glucuronide, a validated urine biomarker for alcohol consumption [33, 34] was depleted. The reduced presence of metformin in the intervention group at 12 months provides a useful positive control as well as offering an opportunity to verify medication usage. In the linear regression analysis, the association of omega-3 (n-3) fatty acids with allocation did not attenuate with additional adjustment for weight change. This suggests that the increased levels of these essential fats in participants from the intervention arm are due to dietary changes (sustained at 12 months) and are not directly related to weight loss.
Metabolite profile variation and clinically relevant biomarkers
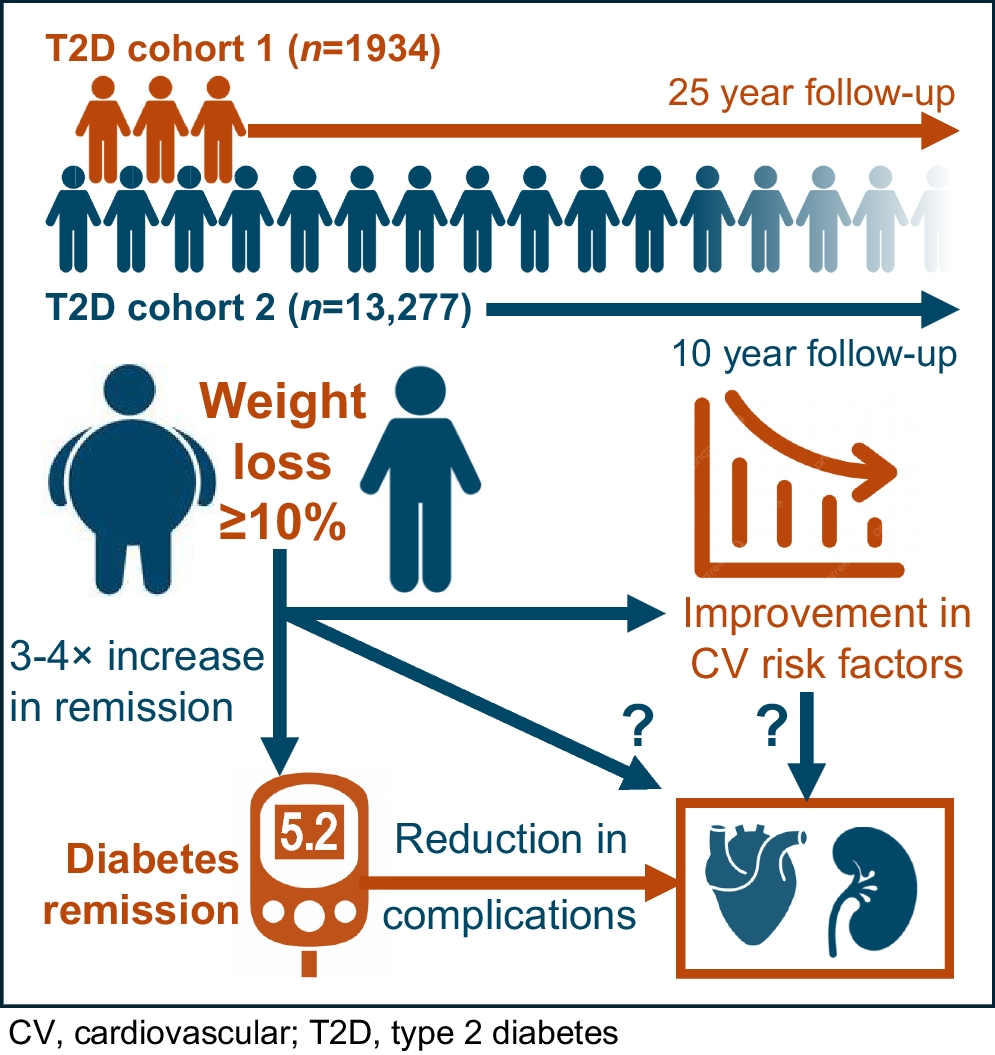
Metabolic profile at 12 months, as captured by the intervention-associated metabolites, was strongly correlated with weight change, possibly explaining much of the allocation effect that we observe. Indeed, the changes we see in levels of glucose and BCAAs are characteristic of those seen with weight change in other settings [20, 35, 36]. We see a decrease in the TG/total lipids ratio across LDLs and VLDLs in participants in the intervention group, with what appears to be a corresponding increase in the total cholesterol and/or cholesteryl ester/total lipids ratio in a similar subset of lipoproteins as would be expected given the previously characterised decrease in hepatic production of VLDL TG following dietary weight loss in type 2 diabetes [37]. These effects are in keeping with the proposed mechanism by which excess TG in the circulation triggers the transfer of TGs from the core of TG-rich lipoproteins to LDL in exchange for cholesteryl esters by the cholesteryl ester transfer protein [38].
The metabolic profile at 12 months was also correlated with change in HbA1c, demonstrating the ability of the intervention-associated metabolites identified to capture changes in glycaemic health as expressed by traditional clinical biomarkers. However, we also found evidence of subtle differences in the metabolome of those who achieved type 2 diabetes remission as compared with those that did not despite similar levels of weight loss. For example, among those individuals who lost the most weight (greater than 9.0 kg) during the trial, those who also achieved diabetes remission showed greater decreases in glucose, fructose and mannose, as compared with those who did not achieve remission. To some extent, this likely reflects the diagnostic criteria on which remission status was based; indeed, 1,5-anhydroglucitol (a proposed marker of short-term glycaemic control [39]) also appears in the list of metabolites that showed differential change by remission status. Based on the current analysis, we are unable to determine whether these differences reflect metabolic processes that contribute to remission or are simply a reflection of an individual’s current metabolic health status. Meanwhile, the correlations observed between intervention-associated changes in metabolites and clinical indicators of liver health suggest that by conducting an in-depth analysis of metabolites in the presence of sustained improvements to liver health, as here, we can further investigate proposed biological systems, such as the twin cycle hypothesis [40], including in the context of variable individual response (see ESM Discussion).



Comments (0)