
Remember me

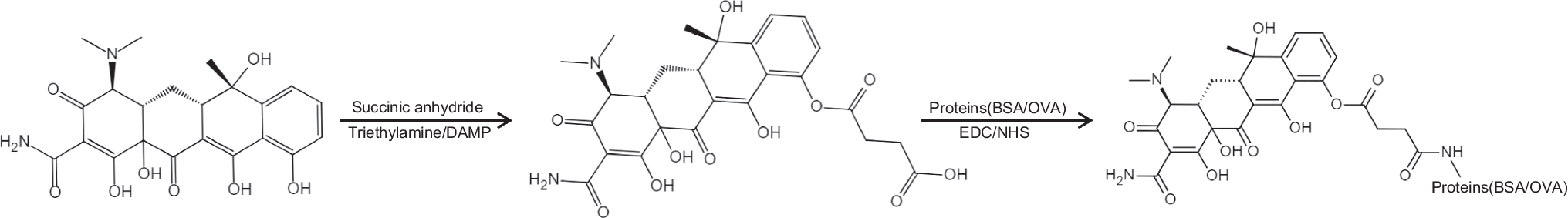
The main objective of the medium optimization was to replace NaCl in the standard medium with other sodium salts in order to prevent chloride-associated corrosion processes in bioreactors.
In the first series of experiments, NaCl was replaced by Na2HPO4 and Na2SO4 in different compositions according to Table 1. In order not to change the supply of sodium ions, the total concentration of sodium ions was kept constant. With these media, cultivations were performed in 500 ml shake flasks. MOPS buffer was added to keep the pH as constant as possible. OD600, glucose concentration, osmolality, and pH were measured at the end of the cultivations. In all cases, glucose was not detectable, indicating that growth was limited by glucose.
Figure 1 shows the results of these experiments. The final OD600 values show that V. natriegens grow similar in all media used. The highest OD600 value of 21.6 ± 1.3 was obtained in medium VN4 with Na2HPO4 without the use of Na2SO4. Medium VN2 with NaCl replaced by Na2SO4 had the lowest OD600 value (18.4 ± 2.3). The results show that NaCl can be replaced completely by other sodium salts.
Fig. 1
Shake flask cultivations of V. natriegens in different media according to Table 1. Values for OD600 (a), osmolality (b), and pH (c) at the end of the cultivation. For each condition, three independent experiments were performed. Error bars show standard deviations
The osmolalities in the new media at the end of the cultivations were between 0.82 and 0.91 Osmol kg−1, which was lower compared to the standard medium VN1 with NaCl (1.08 Osmol kg−1). The final pH values were in the range of 6.1 to 6.8.
The fermentation broth was also visually examined for precipitate formation before and at the end of cultivation. Before the start of the cultivations, no precipitates were visible in the media. In some cases, the samples showed precipitations at the end of the cultivations. Precipitates were clearly visible as a white precipitate below the cell pellet in the centrifuged sample (see Fig. 2).
Fig. 2
Precipitations in the centrifuged sample and microscopic image of precipitated crystals
Microscopic examination of these deposits indicates precipitation of ammonium magnesium phosphate (AMP) due to the characteristic crystals (see Fig. 2). AMP crystallizes in the orthorhombic system as white to yellowish or brownish-white pyramidal coffin-like crystals (Whitaker and Jeffery 1970).
In order to specifically investigate precipitation phenomena in the new media, the influence of pH on medium VN4 (NaCl replaced by Na2HPO4) at room temperature of 20 °C was first examined without cells. Since precipitation can also occur several hours after preparation of the solutions (Pirt 1975), the solution was kept at 20 °C for 24 h after preparation and then tested again for precipitation. The MgSO4 stock solution was added as the last component and just before that, the pH was adjusted to the respective value. Table 4 summarizes the results obtained. No precipitates were observed at pH 6.5. At pH 6.75 and 7.0, no precipitates were visible at the beginning, but precipitates appeared after 1 day. At pH above 7, precipitation occurred immediately after the addition of MgSO4.
Table 4 Precipitations of media VN4 (NaCl replaced by Na2HPO4) depending on the pH valueCultivations in 1 L bioreactor: influence of pH on growth behaviorThe precipitation experiments show that the pH value should be kept as low as possible to avoid precipitation. Therefore, the influence of pH on the growth behavior of V. natriegens was further investigated. Since it is not possible to precisely control the pH in shake flasks, batch cultivations were performed in 1 L Multifors bioreactors at 30 °C and at pH set points of 6.5, 6.75, and 7.0, respectively. For each pH value, two cultivations were performed. To further reduce the potential for precipitation, medium VN3 with a lower phosphate concentration was used, resulting in similar OD yields of 20.5 ± 0.97 compared to medium VN4 with 21.6 ± 1.3 (see Fig. 1). In these experiments, the glucose concentration was increased to 24 g L−1. After 4 h of cultivation, the OD values at pH 6.75 and 7.0 ranged from 33.1 to 37.4, while at pH 6.5, the OD values were much lower at 22 and 24.7, respectively (see Fig. 3).
Fig. 3
Time profiles of OD600 during cultivations of V. natriegens in medium VN3 at different pH values
Therefore, the pH value should be set to 6.75 or above. However, the results above show that precipitation may occur at these pH values.
Further media optimizations using sodium citrateThe results so far show that NaCl can be completely replaced by Na2HPO4 and Na2SO4 with comparable biomass yields. To avoid precipitation in medium VN4, the pH should be adjusted to values below or equal 6.5. However, bacterial growth is reduced at pH values below 6.75. Therefore, it is desirable to further modify the medium to substantially avoid the occurrence of precipitation. This is particularly important for fed-batch cultivations, as critical concentrations for precipitation could be reached by adding media components.
The formation of precipitates can possibly be prevented by the use of complexing agents. For example, EDTA can complex magnesium ions and thereby inhibit the formation of ammonium magnesium phosphate (Das et al. 2017). Citrate can also complex magnesium ions and may contribute to inhibition of AMP formation (Tate et al. 1965; Prywer et al. 2015).
In order to investigate the influence of citrate on the growth behavior of V. natriegens and on the occurrence of precipitates, sodium citrate was used as a further sodium salt in addition to Na2HPO4 and Na2SO4 for elimination of NaCl in the standard medium VN1. For this purpose, another set of experiments with different mixtures of these three sodium salts according to Table 2 (VN5 to VN8) was carried out in shake flask experiments in the SFR at pH 7.0. Again, the total concentration of sodium ions was kept constant in order to have no change in the supply of sodium ions. In another experiment, the medium was supplemented with EDTA instead of sodium citrate (medium VN9 in Table 2). The occurrence of precipitates at the beginning and at the end of the cultivations was examined.
The results in Fig. 4 show that in the EDTA medium, VN9 growth was strongly inhibited. All cultivations with sodium citrate (media VN5 to VN8) grew comparably, with mean OD values ranging from 18.9 to 19.3. In runs with medium VN3, slightly higher OD values of 20.5 ± 0.97 were obtained. The osmolalities at the end of the cultivations were between 0.8 and 0.89 Osmol kg−1. The final pH values were in the range of 6.6 to 6.83 and glucose was not detectable.
Fig. 4
Shake flask cultivations of V. natriegens in different media according to Tables 2 and 3. OD600 (a), osmolality (b), and pH (c) at the end of the cultivation. For each condition, three independent experiments were performed. Error bars show standard deviations
In all media, no precipitations were detected before inoculation and at the end of the runs. Precipitation was practically prevented in all media with sodium citrate. Media VN5 through VN8 were stored sterile without cells at room temperature for another 7 days. Slight precipitation occurred in medium VN5 with the lowest citrate content, while all other media remained precipitation-free. A good compromise between high biomass yield and elimination of precipitation is medium VN6. In a final step to optimize the medium VN6, the existing NH4Cl was replaced by (NH4)2SO4 to further reduce the chloride content. This medium is referred to here as VN10. The composition of the optimized growth media VN6 and VN10 is summarized in Table 3. The OD values obtained in medium VN10 were 20.8 ± 0.87 and therefore slightly higher compared to those of media VN5 to VN8 (see Fig. 4). The media VN6 and VN10 were also used for the fed-batch cultures in the Multifors bioreactors. However, the content of trace elements and the concentration of glucose were doubled (see Table 3).
Sodium citrate was also added to the feed solution as described in section “Materials and methods” and the pH of the feed solution was adjusted to 6.5 to prevent precipitation. No precipitation was observed in the feed during our experiments.
Fed-batch cultivationsFed-batch cultivations were performed in the Multifors bioreactor with the optimized low-chloride medium. With the Multifors, it is possible to operate two bioreactors in parallel. In the first batch (run A), medium VN6 (see Table 3) was used. The temperature was set to 30 °C. Figure 1 and Fig. 4 show that the osmolality of the standard medium VN1 is 1.08 Osmol kg−1 and therefore higher than that of the other media VN2 to VN10 (between 0.8 and 0.91 Osmol kg−1), because the total concentration of osmotically active particles is higher in the standard medium VN1. Therefore, in the second cultivation (run B), medium VN11 was used, in which the concentrations of Na2HPO4, Na2SO4, and sodium citrate were increased (see Table 3 for composition). The aim was to determine whether a higher salt concentration combined with a higher osmolality leads to better growth of the bacteria. These 2 cultivations were repeated with the media VN10 (run C) and VN12 (run D), where NH4Cl was replaced by (NH4)2SO4.
In Fig. 5, the time profiles of cultivation parameters for the fed-batch cultures of V. natriegens (denoted as run A, B, C, and D) are shown. The batch phase lasts approximately 4.2 h for run A and B and 3.5 h for run C and D until glucose was depleted. Biomass concentration increased exponentially while glucose concentration decreased during this phase. The maximum specific growth rate during the batch phase was between 0.9 h−1 in run B and 0.96 h−1 in run D.
Fig. 5
Time profiles of CDW and glucose during high-cell-density cultivations of V. natriegens in the optimized media VN6 (run A), VN11 (run B), VN10 (run C), and VN12 (run D). In run B and D, the sodium concentration was increased (see Table 3). After glucose limitation, the feed solution was fed exponentially indicated by the black vertical line
Figure 6 shows the online parameters DO, stirrer speed, and O2-content in aeration for run C and D (see Supplementary Fig. S1 for the corresponding plots for run A and B). Glucose limitation leads to a sharp increase in DO (see Fig. 6 and Fig. S1). After glucose depletion, the exponential feeding was started with a set point for the specific growth rate of 0.3 h−1. This value was chosen mainly to prevent a limitation in the oxygen supply during the fed-batch phase. To avoid oxygen limitation, the reactor was operated temporarily at the maximum agitation rate of 1200 rpm, and the aeration gas was enriched with pure oxygen (see Fig. 6 and Fig. S1). During the fed-batch phase, the glucose concentration was initially below the detection limit of 90 mg L−1. After 10 h, the glucose concentration began to increase slightly in run C and D (see Fig. 5). The final biomass concentrations achieved in all 4 runs were in the same range (run A: 62.5 g L−1, run B: 62 g L−1, run C: 59 g L−1, run D: 61 g L−1).
Fig. 6
Time profiles of DO, O2-content in aeration, and stirrer speed during high-cell-density cultivations of V. natriegens in the optimized media VN10 (a, run C) and VN12 (b, run D). After glucose limitation, the feed solution was fed exponentially indicated by the black vertical line
During the batch phase, the bacteria produced the by-product acetate (see Fig. 7). After glucose is completely consumed at the end of the batch phase, acetate is used up during the fed-batch phase. At the end of run C and D, its concentration begins to increase again as the glucose concentration increases.
Fig. 7
Time profiles of osmolality (a), ammonia (b), phosphate (c), and acetate (d) during high-cell-density cultivations of V. natriegens in the optimized media VN6 (run A), VN11 (run B), VN10 (run C), and VN12 (run D)
The ammonium concentration in both reactors increases continuously during the batch phase due to the addition of ammonia water by pH control (see Fig. 7). As expected, the osmolality of run A and C was initially lower than that of run B and D. The osmolalities in run B and D are in the range of the osmolality in the standard medium VN1. The growth rate in run B and D decreases towards the end of cultivation, presumably due to temporary oxygen limitation (Fig. 6 and Fig. S1). However, the feed continues to be added exponentially. This causes substrates from the feed (e.g., phosphate) to accumulate in the bioreactor. This in turn leads to an increase in osmolality.
Comments (0)