
Remember me



To investigate potential differences between non-diabetic and diabetic pancreatic cancer, we first screened out DEGs from TCGA database and found 255 up-regulated and 26 down-regulated genes (Figure S1A-B). Enrichment analyses based on GO and KEGG databases showed that the functions of the 281 DEGs were mainly focused on immune signaling pathways, indicating that immune regulation might be an important part of the interaction between pancreatic cancer and DM (Figure S1C-D). The immune signaling pathways mainly included the activation process and internal regulation of B cells and T cells (Figure S1C-D). Furthermore, we performed PPI analysis based on these DEGs and identified seven MCODE complexes (Figure S1E) [17, 18]. Green and red MCODEs represent the signaling pathways of B and T cell receptors, respectively (Figure S1E). The remaining MCODEs were closely related to the migration and functional regulation of immune cells (Figure S1E). These results suggest that re-educating intratumoral immunity may be the major mechanism by which DM influences the carcinogenic process of pancreatic cancer.
To explore the immune characteristics of diabetic pancreatic cancer, we mapped the expression profile to GeneSymbol and performed the deconvolution xCell algorithm to evaluate the abundance of immune cells (Fig. 1A and Figure S2). Compared with samples from non-diabetic group, dendritic cells (DCs), B cells, and CD8+ T cells in samples from diabetic PAAD showed significant increases, while macrophages, natural killer cells (NK cells), CD4+ T cells, basophils, and monocytes showed no obvious differences (Fig. 1A and Figure S2). Meanwhile, diabetic pancreatic cancer exhibited higher immune scores than non-diabetic group (Fig. 1A).
Fig. 1
Immune cell abundance and phenotype undergone significant alterations in diabetic PAAD. A Immune score and abundance of dendritic cells, B cells, and CD8+ T cells by xCell. B Expression level of immune-exhaustion markers in tumor samples of non-diabetic and diabetic PAAD patients. C Pearson correlation analyses between the expression of immune-exhaustion genes and immune cell markers. Pink and red circles indicate R value 0.3–0.8 and > 0.8, respectively. D Single cell type specificity of CD22 and TIGIT and their specific expression in bone marrow and lymph node immune cells from human single cell atlas
Additionally, immune phenotype largely determines the tumor-promoting or tumor-suppressing effects of immune cells [29,30,31]. To further evaluate their phenotypes, we analyzed the RNA levels of several immune markers and found elevated expression of PDCD1, BTLA, EOMES, CD22, CTLA4, TIGIT, and Foxp3 (Fig. 1B). Pearson correlation analysis showed that CD22 was strongly associated with B lineage markers, including CD19, MS4A1, CD79A, and CD79B, while TIGIT was closely related to CD8+ T cell markers with R values more than 0.8 (Fig. 1C). According to the human single cell atlas, CD22 mainly acts on the function of B cells, while TIGIT acts on the immune system and transcription of NK cells and T cells (Fig. 1D). In bone marrow and lymph node samples, CD22 and TIGIT were also expressed at higher levels in B cells and T cells, respectively (Fig. 1D). These results suggested that infiltrating immune cells in diabetic pancreatic cancer might be unable to exert antitumor activity, and CD22+ B cells and TIGIT+ CD8+ T cells were the dominant immune-inhibitory populations.
3.2 Increased distribution of intratumoral immune-inhibitory adaptive immune cellsIn order to determine the unique immune characteristics in diabetic PAAD, multicolor immunofluorescence was carried out on clinical tumor samples. As shown in Fig. 2A–B, the percentages of CD22+ CD19+ B cells and TIGIT+ CD8+ T cells increased significantly in diabetic PAAD, indicating their transition towards immunosuppressive phenotype. Compared with the immune-exclusive condition in the non-diabetic group, more infiltrating CD19+ B cells and CD8+ T cells were observed in the diabetic group (Fig. 2A–B), which was consistent with the results of the xCell deconvolution algorithm.
Fig. 2
The distribution of immune-suppressive adaptive immune cells increased in diabetic PAAD. A The intratumoral distribution of CD22+ CD19+ B cells by multicolor immunofluorescence (n = 10). Scale bar: 50 µm for upper part and 10 µm for lower part. B The intratumoral distribution of TIGIT+ CD8+ T cells by multicolor immunofluorescence (n = 10). Scale bar: 50 µm for upper part and 10 µm for lower part. C-D Body weight and blood glucose of BKS-WT and BKS-DB (n = 3). E-F The phenotype estimation of CD8+ T cells and B cells by flow cytometry (n = 3). All data were expressed as mean ± SD
To simulate the diabetic circumstance, BKS-DB, as a spontaneous diabetes mouse model based on the background of BKS-WT, was adopted in our study [21]. As shown in Fig. 2C–D, higher body weight and blood glucose were detected in BKS-DB mice. Subsequently, subcutaneous tumor model was constructed in BKS-WT and BKS-DB mice, and single cell suspension from tumor samples was obtained to estimate the phenotype of intratumoral adaptive immune cells. As shown in Fig. 2E and Figure S3A, the percentage of CD22+ B cells, which were confirmed to be CD86Low inactive state, significantly elevated in BKS-DB group. Compared with the BKS-WT group, mean TIGIT intensity of CD8+ T cells in BKS-DB group increased by 2.51 times (Fig. 2F). Taken together, diabetic PAAD exhibits an immune-defective landscape, which leads to a poor prognosis and resistance to immunotherapy.
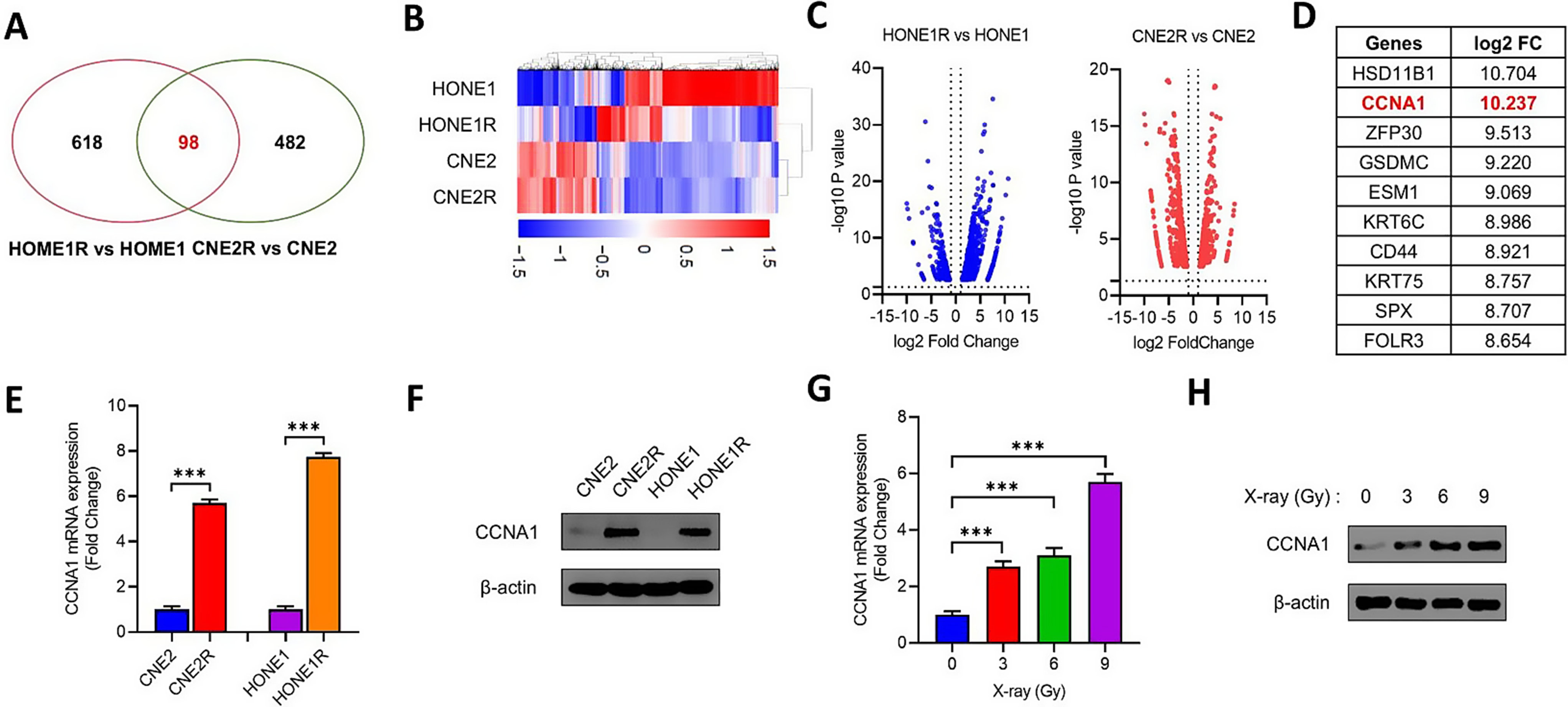
3.3 RNA-sequencing conducted on sorted CAFs from subcutaneous tumorsAccording to spatial transcriptome analysis, B cells and CD8+ T cells are mainly distributed in the stromal compartments of cancers [32,33,34]. CAFs, as the major stromal component, show complicated crosstalk with tumor-infiltrating immune cells. In diabetic PAAD individuals, stroma score significantly increased along with higher immune score (Figure S4A). Considering the multiple associations between fibroblasts and immune cells, we conducted RNA-sequencing on sorted CAFs to identify the involved genes in CAFs that affect immune remodeling in diabetic PAAD. In this part, CAFs were isolated from subcutaneous tumors of BKS-WT and BKS-DB mice and sorted using flow cytometry based on negative selection to exclude cancer, immune, and red blood cells (EpCAM-, CD45-, and TER119-) (Fig. 3A). 3979 genes were found to be differentially expressed in CAFs by at least 1.5 folds, including 1990 up-regulated and 1989 down-regulated genes (Fig. 3B).
Fig. 3
RNA-sequencing conducted on sorted CAFs from subcutaneous tumor. A-B Schematic diagram of RNA-sequencing experiment and volcano plot of DEGs based on RNA-sequencing data (n = 3). C-D Enriched biological pathways of screened DEGs based on GO and KEGG databases. E Enrichment analysis based on Reactome using Metascape. Each cluster was rendered in a different color and marked with p value in the lower part
Enrichment analyses based on GO and KEGG database showed that enriched pathways included cell-cell signaling, inflammatory response, cytokine-cytokine receptor interaction, and TNF signaling pathway (Fig. 3C–D and Figure S5). Meanwhile, enrichment analysis based on Reactome database indicated that cytokine signaling in immune system, as one of the top 20 enriched pathways, may be actively involved in CAF-mediated DM-associated alterations in PAAD (Fig. 3E). These results demonstrated that CAFs could induce immune remodeling in the context of diabetic PAAD.
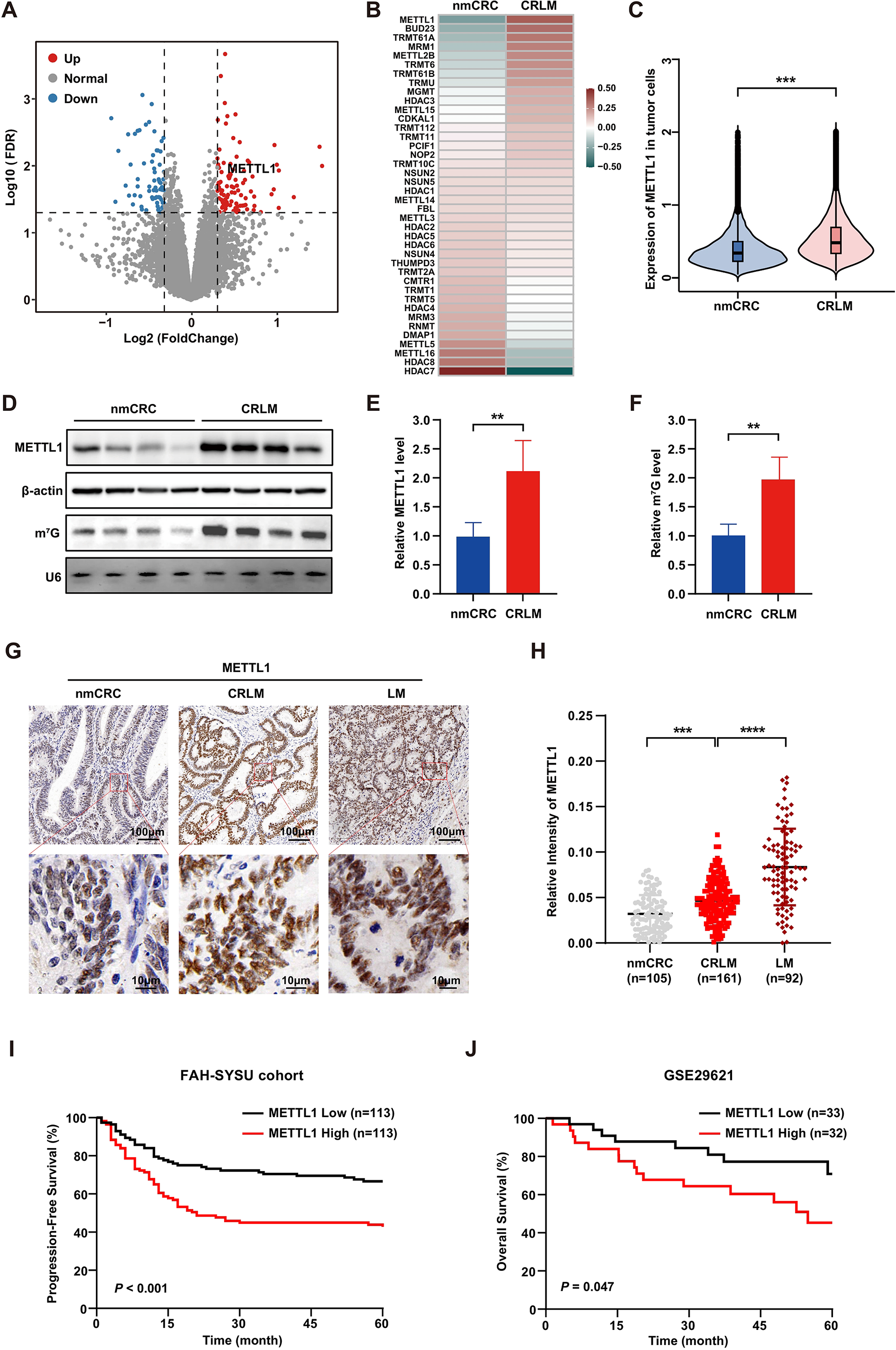
3.4 Overexpressed CXCL12 was associated with immune remodeling in diabetic PAADIn order to investigate the key genes involved in the CAF-mediated DM-associated immune transformation, Venn analysis was performed between DEG sets from TCGA database and our CAF-based RNA-sequencing data, leading to 40 overlapping DEGs (Fig. 4A). Further enrichment and PPI analyses confirmed that the function of these overlapping DEGs was enriched in immune pathways, including cytokine-cytokine receptor interaction and extrafollicular and follicular B cell activation (Fig. 4B–C). These results indicated that immune re-education was the major way in which CAFs induced DM-related alterations in PAAD.
Fig. 4
Overexpressed CXCL12 was associated with immune-suppressive microenvironment in diabetic PAAD. A Venn diagram between DEGs from TCGA database and CAF-based RNA-sequencing data. B-C Enrichment analysis and PPI network based on overlapping genes. D-F Pearson correlation analyses between CXCL12 and immune cell abundance by xCell, Quantiseq, Timer, MCPcounter, and Estimate algorithms
To estimate the abundance of immune subgroups, xCell, Quantiseq, Timer, MCPcounter, and Estimate algorithms were applied in our study (Fig. 4D–F). Among the 40 overlapping DEGs, CXCL12 exhibited positive correlation with the abundance of B cells and CD8+ T cells, with the most consistent results across the above algorithms (Fig. 4D–F). Additionally, CXCL12 was positively correlated with immune-inhibitory markers, including CD22 and TIGIT (Figure S6). In terms of overall survival (OS), we found that higher CXCL12 expression in diabetic PAAD individuals was associated with shortened median OS (Figure S7). Conclusively, these results implied that CXCL12 expressed by CAFs may play a dominant role in DM-induced immune remodeling, potentially leading to poor prognosis.
DM is characterized by activated AGE-RAGE, insulin-like growth factor-1 (IGF-1)-IGF-1 R pathway, hyperinsulinemia, and excessive ROS. Levels of AGEs, IGF-1, insulin, and ROS were firstly examined to explore their roles in DM-induced CXCL12 expression. Compared with BKS-WT, intratumoral AGEs, IGF-1, and blood insulin level showed significant increases in BKS-DB group (Figure S8A). The ROS in CAFs isolated from subcutaneous tumors of BKS-DB mice was 2.63 times higher than that from the BKS-WT group (Figure S8B). When treated with exogenous AGEs, IGF-1, and H2O2, the CXCL12 level of mPSC significantly increased, while the addition of insulin did not induce changes in CXCL12 expression (Figure S8C). FPS-ZM1, JB-1, and NAC were adopted as the inhibitors in the following experiments. As shown in Figure S8C, the induction of CXCL12 by AGEs, IGF-1, and H2O2 was reversed by FPS-ZM1, JB-1, and NAC, respectively. These results demonstrated that AGE-RAGE, IGF-1-IGF-1 R pathway, and ROS were involved in the regulation of CXCL12.
3.5 Elevated distribution of CXCL12High CAFs in diabetic PAADTo confirm the presence of CXCL12High CAFs in diabetic PAAD, multicolor immunofluorescence was performed on tumor samples from PAAD patients with or without DM. As shown in Fig. 5A, the stroma in diabetic PAAD labeled by Desmin was thicker than that in non-diabetic patients. Compared with the non-diabetic group, CXCL12 intensity of each Desmin+ cell was significantly increased in the diabetic group (Fig. 5A). These results demonstrated the increased distribution of CXCL12High CAFs in diabetic PAAD, and indicated that the excessive growth of CAFs and their individual overexpression of CXCL12 both contributed to higher intratumoral concentration of CXCL12.
Fig. 5
CAFs overexpressed CXCL12 in diabetic PAAD. A Representative images of Desmin and CXCL12 immunofluorescence staining on clinical PAAD samples (n = 10). Scale bar: 50 µm for upper part and 20 µm for lower part. B Intratumoral and CAF-expressed CXCL12 concentrations in BKS-WT and BKS-DB animal models (n = 3). All data were expressed as mean ± SD
Meanwhile, subcutaneous tumor model was constructed in BKS-WT and BKS-DB mice, and intratumoral CXCL12 concentration was examined by Elisa. As shown in Fig. 5B, the internal CXCL12 concentration of tumor samples in BKS-DB group was 1.63 times higher than that in BKS-WT group. Compared with BKS-WT group, cytoplasmic CXCL12 of CAFs sorted from BKS-DB group showed a significant elevation by 2.70 times (Fig. 5B).
3.6 CXCL12 promoted immune-inhibitory phenotype of B cells and CD8+ T cells in vitroAdaptive immune dysfunction is commonly present in PAAD and mediates resistance to treatment [35]. As a representative marker of B cells, CD22 can inhibit immune activation during antigen presentation and help malignancy evade immune attack. Multiple studies have demonstrated that increased TIGIT+ CD8+ T cells were closely associated with immune evasion in PAAD, colon cancer, and ovarian cancer [36,37,38]. Considering the phenotypic alterations of B and CD8+ T cells in diabetic PAAD and their relation with CXCL12 expression, in vitro experiments were carried out to confirm the role of CXCL12 in regulating antitumor immunity (Fig. 6). We firstly isolated primary mPSCs and performed IHC detection of α-SMA and Oil Red staining to validate their identity (Figure S4B). Isolated mPSCs were subsequently immortalized by transfection with SV40 large T antigen and telomerase. Primary B cells and CD8+ T cells with a purity of more than 90% were harvested, and co-cultured with mPSCs (Fig. 6A–B), which were transfected with negative control or CXCL12 overexpression plasmids. Tumor supernatant was added to mimic the tumor microenvironment, and plerixafor (500 ng ml−1) was applied as an antagonist of the CXCL12–CXCR4 axis. As shown in Fig. 6A, mPSCoe-CXCL12 co-culture significantly elevated the percentage of CD22+ B cells, while plerixafor treatment reversed the process. Compared with the control group, mean TIGIT intensity of CD8+ T cells in the mPSCoe-CXCL12 group increased by 6.55 times, and subsequently decreased by plerixafor (Fig. 6B). These results implied the crucial role of CXCL12–CXCR4 axis in immune remodeling of diabetic PAAD.
Fig. 6
CXCL12 promoted immune-inhibitory phenotype of B cells and CD8+ T cells in vitro. A Isolation and purification of primary B cells and their phenotype transition when co-cultured with mPSCoe-NC, mPSCoe-CXCL12, or mPSCoe-CXCL12+plerixafor (n = 3). B Isolation and purification of CD8+ T cells and their phenotype transition when co-cultured with mPSCoe-NC, mPSCoe-CXCL12, or mPSCoe-CXCL12+plerixafor (n = 3). C-D Apoptosis and CFSE-based proliferation assay of CD8+ T cells in mPSCoe-NC, mPSCoe-CXCL12, and mPSCoe-CXCL12+plerixafor groups (n = 3). All data were expressed as mean ± SD
Antitumor immunity mediated by CD8+ T cells depends on their quality and quantity. To elucidate the mechanism by which CAF-derived CXCL12 affects CD8+ T cell function, apoptosis assay and CFSE-based proliferation experiment were applied in our study. The co-culture with mPSCoe-CXCL12 induced a significant increase in the apoptotic events of CD8+ T cells by 2.37 times, while the antagonistic effect of plerixafor in turn led to a reduced apoptotic rate by 1.88 times (Fig. 6C). Compared with control group, the proportion of CFSELow CD8+ T cells significantly decreased in the mPSCoe-CXCL12 group, reflecting that CXCL12 retarded the process of T cell proliferation (Fig. 6D). These experiments suggested that CXCL12 could restrain the antitumor capacity of CD8+ T cells by affecting their proliferation and apoptosis.
3.7 CXCL12 exacerbated adaptive immune migration and dysfunction in vivoTo investigate the role of CAF-derived CXCL12 in re-educating tumor immune microenvironment of diabetic PAAD in vivo, we established subcutaneous models by injecting a mixture of mPSCs and Panc02 with a one-to-four ratio. mPSCs were transfected with negative control or CXCL12 overexpression plasmids, and plerixafor was intravenously injected to block the CXCL12–CXCR4 axis (Fig. 7A). As shown in Fig. 7A–C, the tumor volume in mPSCoe-CXCL12 group was significantly increased compared with the limited tumor growth in mPSCoe-NC group. Plerixafor treatment subsequently slowed down and reversed the invasive expansion of tumors in mPSCoe-CXCL12 group, demonstrating the antitumor potential of CXCL12–CXCR4 antagonist (Fig. 7A–C).
Fig. 7
Overexpressed CXCL12 exacerbated in vivo adaptive immune migration and dysfunction in diabetic PAAD. A-C Schematic diagram of animal experiment and harvested subcutaneous tumors from different groups (n = 6). D-E Estimation of intratumoral CD19+ B cells and CD8+ T cells distribution by IHC (n = 6). Scale bar: 50 µm. F CD22+ percentage of CD19+ B cells by flow cytometry (n = 5). G TIGIT expression of intratumoral CD8+ T cells in mPSCoe-NC, mPSCoe-CXCL12, and mPSCoe-CXCL12+plerixafor groups (n = 5). All data were expressed as mean ± SD
After harvesting the tumor samples, IHC was performed to estimate the distribution of B cells and CD8+ T cells. Increased B cells and CD8+ T cells were observed in mPSCoe-CXCL12 group, and this phenomenon was inhibited by plerixafor treatment (Fig. 7D–E). Additionally, we digested the tumor tissue into single cell suspension and assessed the phenotype of immune cells by flow cytometry. The proportions of CD22+ CD19+ B and TIGIT+ CD8+ T cells in mPSCoe-CXCL12 group were 6.46 and 2.66 times higher than those in mPSCoe-NC group, and 3.19 and 2.09 times lower than those in mPSCoe-CXCL12 +plerixafor group, respectively (Fig. 7F–G). These results demonstrated that CXCL12–CXCR4 axis could induce immunosuppressive phenotype, despite its capacity of attracting intratumoral infiltration of immune cells. In conclusion, CXCL12–CXCR4 axis, as a classical chemotaxis signaling pathway, was the dominant mechanism regulating DM-related alterations in intratumoral immune infiltration and function.
In order to estimate the immune phenotype of intratumoral B cells, PD-L2, IL-10, and IL-35 were tested. As shown in Figure S3B-C, there was no significant change in the abundance of PD-L2+ B cells between BKS-WT and BKS-DB groups, and intratumoral PD-L2+ B cells exhibited CD22- expression pattern. No upregulated abundance of memory B cells suggested the inactive state of infiltrating B cells. Interestingly, IL-10 and IL-35, which were known to be the markers of B regulatory cell (Breg), showed significant increases in the tumor homogenate from BKS-DB mice (Figure S3D). Higher IL-10 and IL-35 were detected in the co-culture system with mPSCoe-CXCL12, which could be inhibited by plerixafor (Figure S3E). These results indicated that Breg may play a role in the DM-mediated immune remodeling.
Comments (0)