
Remember me



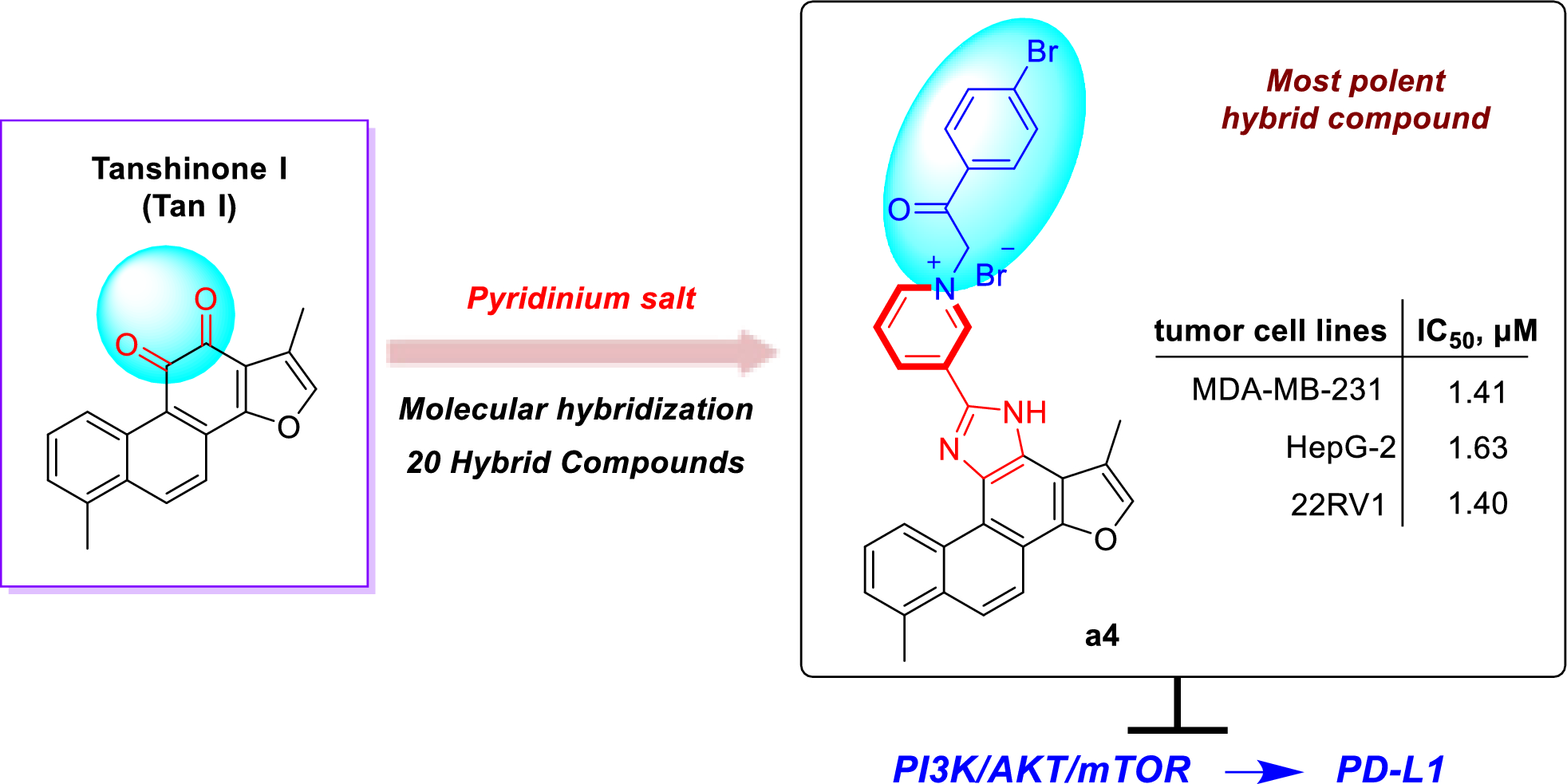

The examined isolates were assigned to species based on sequence analysis of partial tef1α sequences ( provided in the supplementary file). Figure 1 shows the phylogenetic relation between the strains investigated in this article and the type strains of closely related species. T. koningii SZMC 28387 and T. atroviride SZMC 28748 showed 100% identity with their type strains. Strain SZMC 28747 originally was identified as T. fertile, however, it showed 100% identity with the type strains of T. hamatum. T. cf. strigosellum SZMC 28391 and SZMC 28007 could not be identified to species level, therefore, cf. strigosellum was introduced. T. taiwanense SZMC 28390 and T. cf. dorothopsis SZMC 28005 did not match with their type strains either, but were similar to T. dorothopsis type strain and were named as T. cf. dorothopsis SZMC 28390 and T. cf. dorothopsis SZMC 28005.
Fig. 1
Phylogenetic tree of the examined strains from clade Viride based on their partial tef1α sequences
3.2 Peptaibiome of the investigated Trichoderma strainsOut of the 7 strains investigated, (Table 1), 5 produced both peptaibols and lipopeptaibols (T. cf. strigosellum SZMC 28391 and SZMC 28007, T. koningii SZMC 28387, T. hamatum SZMC 28747, and T. cf. dorothopsis SZMC 28005), while T. cf. dorothopsis SZMC 28390 and T. atroviride SZMC 28748 did not produce any lipopeptaibols, their peptaibiome consisted of exclusively SF1 peptaibols (Table 2, Supplementary Fig. 1). The two T. cf. strigosellum strains produced ∼40/60% ratio of peptaibols/lipopeptaibols T. koningii SZMC 28387 and T. cf. dorothopsis SZMC 28005 produced one third ratio of peptaibols of the peptaibiome, while T. hamatum SZMC 28747 produced only one fourth of peptaibols of the peptaibiome (Table 2). Based on their length, lipopeptaibols could be categorised into the short (6–7-residue long), medium (10–11- residue long) or long (15-residue long) groups. The most amount of short lipopeptaibols was produced by T. cf. strigosellum SZMC 28007 with 61.35% of the peptaibiome, while the greatest number of long lipopeptaibols were produced by T. hamatum SZMC 28747 with 40.36% of the peptaibiome. The amount of medium lipopeptaibols was ranging from 30 to 65% of the total production (Table 2).
Table 2 Ratio of peptaibiotics (peptaibiome) produced by the members of clade Viride investigated in this study3.3 Peptaibol profiles of the investigated Trichoderma strainsTables 3 and 4 were obtained from the chromatographic and MS analysis of the investigated Trichoderma strains and show the subfamily 1 (SF-1) peptaibols of the investigated Trichoderma strains. The diagnostic fragment ions resulted by MS2 fragmentation are collected in Supplementary Table 1.
Table 3 SF1 peptaibols produced by Trichoderma strains from clade VirideTable 4 Similarities of SF1 peptaibols produced by Trichoderma strains from clade Viride to known peptaibols3.3.1 Identification of new Strigaibol sequences and the new groups of Strigosellins from T. cf. strigosellumInvestigation of the peptaibol profile of T. cf. strigosellum SZMC 28391 revealed twenty-one 19-residue peptaibols belonging to the SF-1 peptaibol family (Tables 3 and 4). Appart from the known Strigaibol sequences, 14 proved to be new. Their closest positional isomers are Strigaibol sequences described by Park et al. [33]. MS and chromatographical analysis of T. cf. strigosellum SZMC 28007 revealed the production of 14 peptaibols novel to science and were named as Strigosellin A as well as further 6 sequences named as Strigosellin B peptaibols. These peptaibols showed a great similarity to the sequences of Trichokonin TKO-V, Trichokonin V, Tricholongin B and Tricholongin LBI sequences [34,35,36,37].
Strain T. cf. strigosellum SZMC 28391 produced two peptaibols in higher quantities, which were Strigaibol XII and XIV (36.15% and 27.59% of the total peptaibol production, respectively). Four more sequences were produced above 4%, Strigaibol V, VI, X and XX (5.14%, 6.44%, 4.34% and 7.2%, respectively). Strain T. cf. strigosellum SZMC 28007 produced Strigosellin A IX covering 14.55% of the total peptaibol production, while Strigosellins A XII, XIII, and XIV were also produced in relatively higher quantities (10.73%, 13.01% and 10.95%, respectively). Strigosellins A II, V, VI, and VIII, were also produced in significant quantities between 2.78 and 7.41%, while Strigosellins A I, III, IV, VII, X, and XI were produced at 0.48–1.78% of the total peptaibol production. Strigosellins B I, II and V were produced also in significant quantities (5.38%, 8.2% and 6.51%, respectively), while Strigosellin B I, B III, B IV and B VI were below 2.44%.
3.3.2 New Trikoningin KA-like peptaibol compounds from T. koningiiT. koningii SZMC 28387 produced thirty-eight new sequences, while nineteen were positionally isomeric with the previously described Trikoningin KA V from T. koningii described by Goulard et al. [29], and Trikoningin KA-like compounds Ia, IVa, IVb, Va, VIa and XI from T. gamsii SZMC 1656 described by Marik et al. [38] (Tables 3 and 4).
T. koningii SZMC 28387 also produced two peptaibols, Trikoningin KA-like XLIII and XLV (25.83% and 28.48%, respectively) in high quantities, while apart from Trikoningin KA-like XL, XLVI and XLVIII (4.65%, 6.25% and 6.74%, respectively), the rest of the peptaibols were produced for less than 3% and mostly below 1%.
3.3.3 Identification of the new group of Dorothopsins from T. cf. dorothopsisStrain T. cf. dorothopsis SZMC 28390 produced thirteen 19-residue peptaibols also belonging to the SF-1 peptaibol family. All the thirteen compounds were completely new (Tables 3, 4) and their closest positional isomers are Trichostrigocins TSG-A and TSG-B described by Degenkolb et al. [37]. These new compounds were named as Dorothopsins A-a I-XIII (Table 3). However, 18-residue peptaibols, Dorothopsins A-b I-VIII and Dorothopsins A-d I-III, as well as, the 17-residue Dorothopsin A-c I and Dorothopsin A-f I-II sequences, furthermore, a 16-residue Dorothopsin A-e I sequence were also produced by this strain (Table 3). All Dorothopsin A sequences are also new additions to literature (Table 4). Strain T. cf. dorothopsis SZMC 28390 produced Dorothopsin A-a VII covering more than half of the total peptaibol production (54.05%). After that, Dorothopsins A-a VIII and A-d I, were produced the second highest with 10.48% and 8.35%, respectively. Among the Dorothopsin A-a sequences, a few peptaibols were 1.5%–4.5%, but the rest and all Dorothopsin A-b, -c, -d, -e and -f sequences were below 1.5% apart from Dorothopsin A-d I with 8.35%.
T. cf. dorothopsis SZMC 28005 produced thirty-four new sequences, which were the most unique among the already known SF1 peptaibols and named as Dorothopsin B. They had the most varied sequences, sometimes with differences in 6 positions from the already known Tricholongin LBI sequence described by Degenkolb et al. [37]. The most interesting characteristic of these sequences is the substitution of the acetylated Aib1 (Ac-Aib1) and Phe3 known from Tricholongin sequences to the acetylated Phe1 (Ac-Phe1) and Aib3. Furthermore, Phe1 was also substituted with Trp1 in several Dorothopsin B sequences. The most produced sequence by T. cf. dorothopsis SZMC 28005 was Dorothopsin B XXVI with a relatively low quantity of 10.15%, which was followed by Dorothopsins B VII, XIII, XV, XXV, XXXI, and XXXIV (5.05%, 9.71%, 7.95%, 6.16%, 4.01% and 4.53%, respectively). All the other sequences were produced below 4%.
3.3.4 Identification of new Trichorzianin TA sequences and peptaibol-like compounds from T. atrovirideT. atroviride SZMC 28748 produced thirty-three peptaibol compounds, as well as fourty-one incomplete peptaibol sequences named as Peptaibol-like compounds in Tables 3 and 4. The thirty-three peptaibol sequences were positionally isomeric with the Trichorzianin TA sequences [41], out of which, thirteen proved to be new. The Peptaibol-like sequences were also identical with the thirty-three trichorzianin TA sequences [41], but unusual at their C-terminus, where the last part of the sequences is shown with the remaining mass difference, which were Δm = 145, 146, 160, or 222 D, furthermore, all MS2 spectra contain the water loss [y6–H2O]+ fragment ions (Supplementary Fig. 2), which usually occurs when the C-terminus is an aminoalcohol. The sequences contain only two variable positions, R5 contains Aib/Vxx, while R14 Vxx/Lxx. All other residues are the same in the sequences, compared to each other. These sequences were produced in a relatively low concentration (all below 0.25%), therefore, they were not investigated further.
The two most produced peptaibols of T. atroviride SZMC 28748 were Trichorzianin TA-like XXV and XXIX with 15.24% and 17.59% of the total peptaibol production. They were followed by Trichorzianin TA-like VII, XV, XX, XXII, XXVII and XXVIII peptaibols with 7.68%, 8.99%, 5.26%, 5.44%, 6.19% and 5.34%, respectively. The rest of the peptaibols produced by this strain were around or below 1–2%. All the 41 Peptaibol-like compounds produced by T. atroviride SZMC 28748 were below 0.25% of the total peptaibiome.
3.3.5 Identification of new Tricholongin sequences from T. hamatumThe chromatographic and MS analysis of the T. hamatum SZMC 28747 extract revealed the production of twenty-one 19-residue-peptaibols belonging to the SF-1 peptaibol family (Tables 3 and 4). Among them, 6 peptaibols were new to science and the other sequences were identical to already determined sequences of Tricholongins LB I, II, III, and IV [37]. The newly identified sequences also shared similarities with the sequences of Tricholongins LB I, II, III, and IV. The known peptaibols are produced by T. cf. strigosum BBA 69577 and T. cf. pubescens BBA 66989 strains [37].
Tricholongin LB-like XIII and XVI sequences produced by T. hamatum SZMC 28747 covered 25.68% and 29.09% of the total peptaibol production, which were followed by Tricholongin LB-like XVII and XIX sequences (7.13% and 10.03%, respectively). All other sequences were produced mostly around 1%.
3.3.6 Module skipping hypothesisThe microheterogeneity of non-ribosomal peptides like peptaibols arises from the ability of NRPSs to incorporate various amino acids [42]. Usually, Trichoderma species produce 1 to 4 main peptaibol compounds in high quantities, while the rest of the peptaibol variants are in much lower quantities, which can be the result of the modular synthesis and the misassembly of the peptaibol compounds by NRPS and the lack of repair mechanisms compared to ribosomal synthesis. This may be the reason for the wide range of new peptaibols that can be identified even from already known peptaibol subgroups. This hypothesis needs a thorough genetic data support based on the number of NRPS genes that can be found in the available Trichoderma full genomes.
Based on the results of this study, we hypothesize that the 16, 17 and 18-residue peptaibols produced by T. cf. dorothopsis SZMC 28390 also occured through a ‘module skipping’ event, which means that the NRPS enzyme skipped a module during the synthesis [27, 39]. Two skipping mechanisms were proposed in the literature: direct intermediate transfer between nonadjacent modules caused by a “loss of function mutation” [40], or nonfunctional thiolation or condensation domains of the NRPSs during biosynthesis [39]. Module skipping is usually seen in the case of 10-, 13-, 18-, and 19-residue peptaibols. Similar results were found in the case of SF-1 peptaibols produced by T. flagellatum, T. sinense and T. parareesei, which produced 19-residue peptaibols among the 20-residue peptaibols, but the investigation of the genome of the producer strains revealed no extra 19-module NRPS synthetases, only the gene of a 20-module enzyme was present [27]. Based on this hypothesis, the positions at which module skipping most probably occurred are shown in Table 4 with a minus sign. Our prediction is based on comparing the sequences of 16-residue (Dorothopsin A-e), 17-residue (Dorothopsin A-c) and 18-residue (Dorothopsins A-b and A-d) peptaibols to the sequences of 19-residue peptaibols (Dorothopsin A-a) (Tables 3 and 5).
Table 5 The 1most produced sequences, 2consensus sequences and 3amino acid varieties of the peptaibols produced by each strain3.4 Comparative analysis of peptaibols produced by species of clade VirideThe microheterogeneity within the sequences occurs in the positions where one amino acid is substituted by another one. The strictest positions were found to be in the middle of the sequences usually appearing as the Gln6-Aib7-Aib8-Aib9-Ser10-Lxx11-Aib12-Pro13-Vxx14 motif. This motif was different only in Strigosellin A sequences, where it was substituted with Gln6-Aib7-Aib8-Ala9-Ala10-Vxx11-Aib12-Pro-13-Phe14/Lxx14 produced by T. cf. strigosellum SZMC 28007. R1 usually contained an acetylated Aib except for a few T. cf. strigosellum SZMC 28391 and T. cf. dorothopsis SZMC 28390 sequences, where it was often substituted with Vxx. Acetylated Vxx is not present in SF-1 peptaibols, only in 11–16-residue peptaibols [10, 34, 37, 39]. Mostly, AcAib can be found in SF-1 peptaibols, however, several compounds produced by these strains contained another amino acid residue at the N-terminus (Table 3). Dorothopsin B sequences produced by T. cf. dorothopsis SZMC 28005 always contained Phe1 or Trp1. Boletusin 1, Chrysospermins and Peptaivirins, a group of another 19 residue-peptaibols identified from Boletus spp., Apiocrea chrysosperma and Trichoderma spp. (KGT142), respectively, also contained AcPhe at their N-termini [43,44,45]. Phe occurred strictly in R1, R3, R14 and R19 positions in T. cf. dorothopsis SZMC 28005, T. hamatum SZMC 28747, T. cf. strigosellum SZMC 28007 and T. atroviride SZMC 28748, respectively, while the other strains investigated did not contain Phe at all. The difference in Strigosellin A and B sequences relies in Phe14 and Lxx14 from T. cf. strigosellum SZMC 28007. Interestingly, both types are produced almost equally by this strain. Regularly, Phe is close to one of the termini of the sequences in all peptaibols, however, Phe is in the middle of the sequences in Strigosellin A peptaibols. This strain shows production of both Strigosellin A (Phe14) and B (Lxx14) sequences, as if the production of the regular Lxx14 containing peptaibols were replaced to the rare Phe14 sequences. AcPhe1 was often substituted with AcTrp1 in Dorothopsin sequences, also observed in the literature for 15–16-residue peptaibols (Ampullosporins and Zervamicins) produced by Sepedonium ampullosporum (HKI-0053) and Emericellopsis salmosynnemata, respectively [46, 47]. Sequences of T. hamatum SZMC 28747 contained Ala1 instead of Aib1 in case of only 2 sequences. Only three 20-residue sequences (Trichoaureocin 1a, Trichokonin IIb and Atroviridin D) with AcAla were identified from T. aureoviride, T. koningii Oudemans and an unidentified Trichoderma sp., respectively [35, 48, 49], out of which, the first two species belong to clade Viride, while the latter one was later reidentified as T. arundinaceum belonging to clade Brevicompactum [50]. Gly2 was substituted to Ala or Ser in several sequences of all strains investigated, while R3 contained mostly Ala or Aib. However, in the sequences of T. hamatum SZMC 28747, Phe or Tyr was always identified at the R3 position. Tyr has never been identified from peptaibols at this position, only from lipopeptaibols and other peptaibiotics, such as LP237 F5 and Trichoatrokontins, respectively [18, 51, 52], as well as from all-Aib-replaced peptides by Amycolatopsis azurea, used for bioconversion of AS 1387392 [53]. R4 was strictly Aib in all sequences except for a few sequences produced by T. hamatum SZMC 28747. At R5, mostly Vxx and Lxx were identified, except for T. hamatum SZMC 28747 and T. cf. strigosellum SZMC 28007 sequences, which mostly contained Aib5. This position was followed by the R6-R7-R8-R9-R10-R11 and R12-R13-R14 motifs discussed above. The module skipping occurred at R9 position in all Dorothopsin A-b, -c and -e sequences. R15 contained mostly Aib like in R4, except for a few sequences produced by T. cf. strigosellum SZMC 28007, where it was substituted to Vxx in several sequences. R16 contained either Ala, Aib, Vxx or Lxx which determined the mass of the y7 ion fragments. Gln16 was often substituted with Glu which, apart from the mass of the y7 ion fragments, defines the acidity of the peptaibols as Gln is neutral and Glu shows acidic characteristics. Dorothopsin A sequences also showed the 2nd module skipping at this position. One Dorothopsin A sequence (Dorothopsin A-e I from Table 3) showed in total 3 module skippings at R9, R16 and R17. R18 mostly contained Gln except for Dorothopsin A compounds, where it was often substituted with Gly or Ala. The aminoalcohol was mostly Vxxol or Lxxol, except for Dorothopsin A and Tricholongin LB-like sequences, which often contained Alaninol (Alaol) or phenylalaninol (Pheol)/tryptophanol (Trpol) at the C-terminus, respectively. Pheol is often identified as the C-terminus of peptaibols. The first naming of peptaibols by Pandey et al. [54] as ‘peptaibophols’ was also based on the inclusion of Pheol, which was defined as peptide antibiotics containing the marker amino acid Aib and a C-terminal Pheol residue [54, 55]. After this, additional 2-alcohols such as valaninol (Valol) in TXT-A40 and Leuol in Hypelcin from Hypocrea peltata (current name: T. peltatum) were detected as C-terminal constituents, the above definition of ‘peptaibophols’ needed to be revised and updated. Nevertheless, Trpol is rare at the C-terminus of peptaibols. It was only identified from Boletusin 1, Chrysospermins, Peptaivirin A, as well as Trichorzianin TA and PA sequences [41, 43,44,45, 56, 57].
In a few cases, the consensus sequence (which is a created sequence based on the most abundant amino acid present at each position) was different from the most produced sequence of the strain. These residues are underlined in Table 5. Trikoningin KA-like peptaibols mostly contained Gln at R17 while the most produced Trikoningin KA-like XLV had Glu at this position. Exactly this could be observed among Dorothopsin B sequences where Dorothopsin B XXVI contained Glu instead of Gln. Similarly, Trichorzianin TA-like peptaibols at R14 contained Lxx instead of Vxx in the most produced Trichorzianin TA-like XXIX. Tricholongin LB-like sequences equally produced Aib and Vxx containing sequences at R16, however, the most produced Tricholongin LB-like XVI had Vxx at this position.
3.5 Lipopeptaibol production of the investigated Trichoderma strainsStrain T. cf. dorothopsis SZMC 28390 and T. atroviride SZMC 28748 did not produce any lipopeptaibols (Table 1), their peptaibiomes consist of exclusively SF1 peptaibols. In Tables 6 and 7 the sequences are ordered based on their similarities to each other, the producer strains are shown in the 2nd column. The diagnostic fragment ions resulted by MS2 fragmentation are collected in Supplementary Table 2.
Table 6 Lipopeptaibols produced by Trichoderma strains from clade Viride The area percentages are calculated based on the total lipopeptaibol production of each strainTable 7 Similarities to known compounds of lipopeptaibols produced by Trichoderma strains from clade Viride3.5.1 Identification of the new group of Lipostrigosellins from T. cf. strigosellumT. cf. strigosellum SZMC 28391 produced a total of 12 lipopeptaibols, out of which, four were 6-residue long, while eight were 7-residue long (Table 6). Seven sequences were new, while five were positionally isomeric with already identified Lipostrigocin LSG sequences [37] (Table 7). All the 6-residue lipopeptaibols were new to science and named as Lipostrigosellin I-IV. They are different from 7-residue lipopeptaibols in skipping of Aib/Vxx4. Other Trichoderma species were not found to produce 6-residue lipopeptaibols. Only Halovir A-E sequences from Scytalidium sp. CNL240 [58] and Lipohexin from Moeszia lindtneri HKI-0054 [59] were identified as 6-residue lipopeptaibols. Strain T. cf. strigosellum SZMC 28391 also produced a unique compound in a relatively high amount, the amino acid composition of which could not be revealed. The mass differences (Δm) on the MS2 spectra are shown in Table 6, which is a repetition of 72 and 127 mass differences. Strain T. cf. strigosellum SZMC 28007 produced nine already known and seven new 7-residue sequences which were named as Lipostrigaibols I-XVI. The sequences always started with an OcGly only known from Lipostrigaibols [33]. It is followed by an Ala2, which is again unique, because this position is usually taken by Gly2. R3 and 4 were highly variable, Ala/Aib/Vxx/Lxx and Ala/Aib/Vxx were at these positions. R5 was strictly Ala and R6-7 contained either Vxx-Lxxol, or Lxx-Lxxol, which are usual in the lipopeptaibols (Table 7).
Strain T. cf. strigosellum SZMC 28391 produced two lipopeptaibols in higher quantities, which were Lipostrigocin LSG-like VIIIb and VI, with 37.08% and 31.96% of the total lipopeptaibol production, respectively (Table 6). The “unidentified compound” mentioned above was also produced in a relatively higher amount, 17.45%, compared to the rest of the lipopeptaibols, which were all below 4% (Table 6). The 7-residue lipopetaibols were more produced than the 6-residue lipopeptaibols, the latter all being below 2.17%. Three lipopeptaibols were produced in high quantities by T. cf. strigosellum SZMC 28007, Lipostrigaibols XV, XIII and XVI (23.7%, 22.15% and 20.8%, respectively) followed by Lipostrigaibols XIV and XII with 9.97% and 8.77%, respectively. The rest of the sequences were all produced below 5.22%.
3.5.2 New Lipostrigocin LSG-like sequences peptaibol compounds from T. koningiiT. koningii SZMC 28387 produced a total of 25 lipopeptaibols, out of which seven were 7-residue long while eighteen were 11-residue long (Table 6). Twelve sequences were new, while thirteen were positionally isomeric with already identified Lipostrigocin LSG sequences [37].
T. koningii SZMC 28387 produced more 11-residue than 7-residue lipopeptaibols, out of which Lipostrigocin LSG-like XIV was the most produced (37.46%). Lipostrigocin LSG-like XV, XVI and XX were also produced in a relatively high amount, at 9.6%, 11.01% and 11.37%, respectively. The 7-residue lipopeptaibols were produced below 1.28%.
3.5.3 Identification of the new group of Lipohamatins from T. hamatumT. hamatum SZMC 28747 produced sixty-two 7-, 10-, 11- and 15-residue lipopeptaibols, out of which 16 compounds were known, while 47 were newly identified, out of which all the 10-residue lipopeptaibols were completely new to science and were named as Lipohamatin I-IV. They were similar to Linopubescin LPB A, B and C [37]. The 7-, and 11- residue sequences showed similarity to the Lipostrigocin LSG sequences, while the 15-residue compounds to Trichogin GB IX [37, 60], which was the only 15-residue lipopeptaibol reported.
T. hamatum SZMC 28747 produced the most diverse lipopeptaibol profile. The lipopeptaibol most produced by this strain was Lipostrigocin LSG-like XXXIX with 11.1%. Trichogin GB IX-like XXVI was the most produced 15-residue lipopeptaibol by this strain (6.67%). All 7- and 10-residue lipopeptaibols were produced below 1.71%.
3.5.4 New Lipostrigocin LSG-like peptaibol sequences from T. cf. dorothopsis
Comments (0)