
Remember me
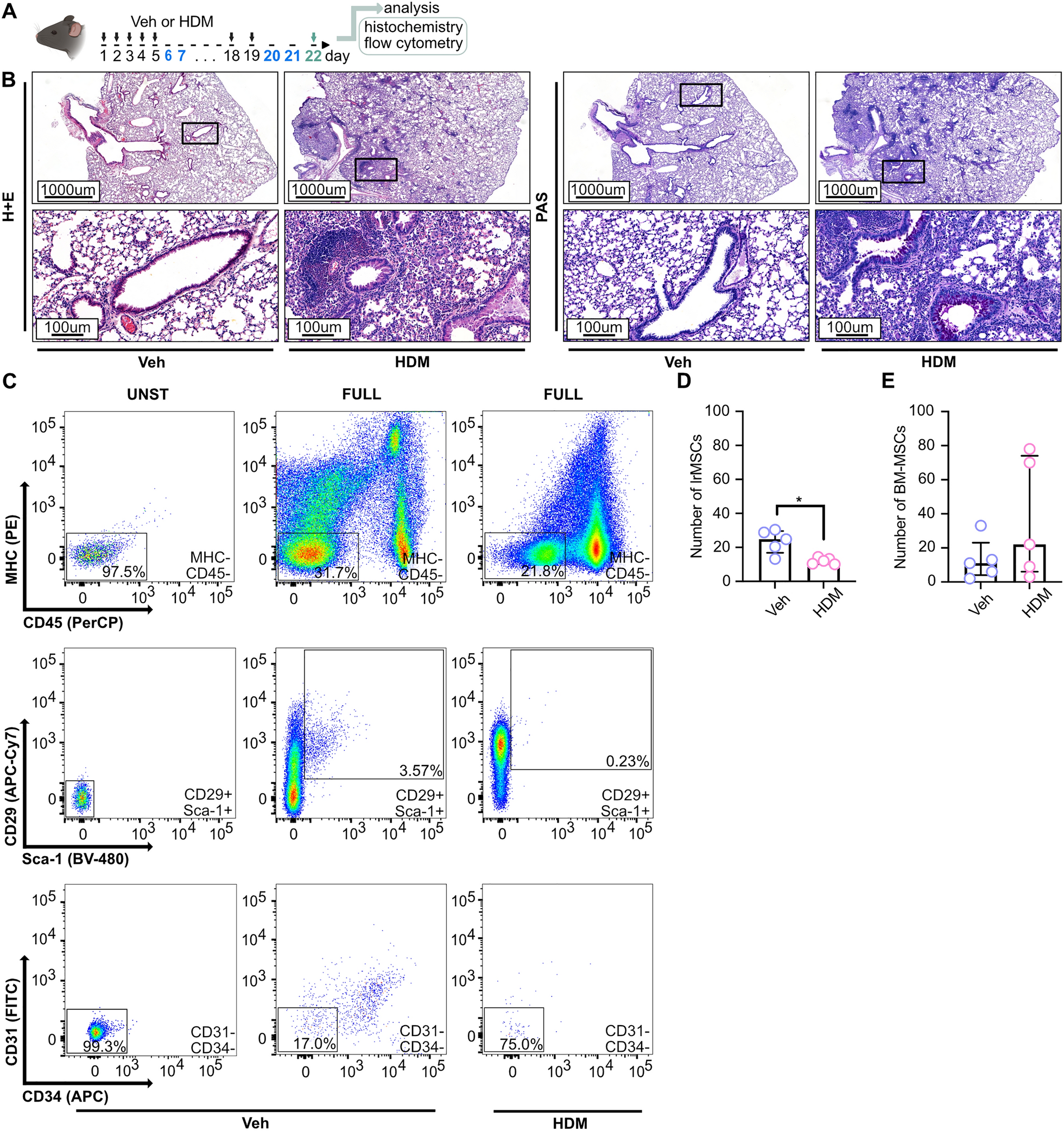


We previously demonstrated that subretinally injected tdTomato-labeled PRPCs, derived from a WA09 CRX+/tdTomato hESC [21] line, survive long term in immunosuppressed normal canine retinas (n = 4 dogs). Donor cells remained detectable for up to 22 weeks post-injection, primarily within the subretinal space, with occasional migration into the host ONL. A subset of cells exhibited cone-like morphology, suggesting early photoreceptor differentiation. In contrast, when PRPCs were transplanted into rcd1/PDE6B dogs at a late stage of degeneration (~ 2 ONL rows remaining), migration was more frequent but disorganized, with cells localizing to both the ONL and inner retina. These donor cells did not adopt photoreceptor-like features, underscoring the negative impact of structural alterations in end stage disease on PRPC integration and maturation [2].
To assess how disease type and/or the stage (degree of ONL loss) at the time of transplantation could influence PRPC fate, we conducted similar experiments in two additional non-allelic canine models at different stages of retinal degeneration. Across all four models -normal, rcd1/PDE6B, RHOT4R/+, and xlpra2/RPGR - we observed substantial loss of tdTomato fluorescence within the first week post-transplantation (Fig. 1). Since the outcomes in normal and rcd1/PDE6B retinas have been previously characterized [2], the current study focuses on the RHOT4R/+ and xlpra2/RPGR models to further evaluate how disease severity influences long-term donor cell survival and integration. First, we assessed the survival of PRPCs following subretinal injection into mutant RHOT4R/+ dogs at end stage retinal degeneration, where only a single row of cone nuclei remained in the ONL. These dogs had previously been acutely exposed to light levels that trigger rapid and complete loss of rods in the central to mid-peripheral retina [24, 25]. Fundus fluorescence photography revealed qualitatively a significant reduction in tdTomato fluorescence within the first week following injection in all 4 injected RHOT4R/+dogs (four eyes). Notably, in contrast to the findings in normal and rcd1/PDE6B dogs [2], further reduction in tdTomato signal was observed in three out of four RHOT4R/+ animals despite uniform immunosuppressive regimen (Fig. 1c–f). A similar continuous decline in tdTomato fluorescence was also noted in one xlpra2/RPGR mutant dog injected at 88 weeks of age, when retinal degeneration was at end-stage disease (~ one row of nuclei remaining in the ONL) (Fig. 1g). To determine whether this fluorescence loss reflected true donor cell clearance, we performed endpoint histological and immunohistochemical analysis in all animals. Staining for human nuclear marker Ku80 along with visualization of tdTomato confirmed complete loss of donor cells in three of the four RHOT4R/+ dogs and in the xlpra2/RPGR dog injected at end stage. In one RHOT4R/+ dog (EM401), a small number of surviving PRPCs were detected localized to the subretinal space (Supplementary Fig. 1A–E). These results suggest that the retinal environment in end-stage disease may not be conducive to the long-term survival of transplanted cells.
Fig. 1
Retinal imaging following subretinal transplantation of hESC-CRX+/tdTomato-derived photoreceptor precursor cells (PRPCs) following subretinal transplantation in canine models of retinal degeneration. Fundus fluorescence images of tdTomato-expressing PRPCs at early and extended time points post-transplantation. a–b Early imaging of grafts in (a) Normal (AS2-427) and (b) rcd1/PDE6B (2299) retina, immediately following subretinal injection and 1 week later. Both animals were included in our previous study (Ripolles-Garcia et al., 2022); however, the images shown here have not been published previously. c–h Longitudinal imaging of tdTomato⁺ grafts in RHOT4R/+ and xlpra2/RPGR dogs at multiple time points following subretinal injection, in animals with end-stage c–g and late-stage (h) retinal degeneration. All animals received systemic immunosuppression. In panel g, the dark shading in the inferior fundus on the day of injection is due to normal pigmentation of the retina and not indicative of hemorrhage; the surrounding gray area corresponds to the retinal detachment caused by the injection bleb. This image was acquired using low flash intensity due to the brightness of the tdTomato signal
To assess whether transplantation at a less advanced stage of photoreceptor degeneration (~ three rows of nuclei left in the ONL or late-stage degeneration) would promote long-term survival of PRPCs, a 43-week-old xlpra2/RPGR dog was injected and followed for up to 22 weeks post-injection (PI). A significant loss of cells occurred within the first week PI, however more tdTomato fluorescence was observed throughout the treated area and was stable on in vivo imaging up to 20 weeks PI, the final imaging timepoint before termination at 22 weeks for histological analysis (Fig. 1h). Upon termination, histological and immunohistochemical assessment confirmed the presence of tdTomato-positive PRPC foci distributed throughout the treated/bleb area. These cells were predominantly localized either within the subretinal space or had migrated into the host retina, though in a disorganized pattern (Supplementary Fig. 1F). Interestingly, at a single locus, PRPCs that had differentiated predominantly into cones were found to be perfectly aligned in the host’s ONL, immediately below the external limiting membrane. These cells formed a short inner segment and extended a short axon with pedicle-like synaptic terminals (Fig. 2). Notably, immunostaining for RPE65 in adjacent sections confirmed that these cells were integrated beneath the ELM and not merely apposed to the RPE, supporting true structural integration into the host retina (Fig. 2C2).
Fig. 2
Differentiation, migration, and synapse formation after transplantation of donor human PRPCs in the xlpra2/RPGR host retina at late-stage disease. Immunohistochemical analysis of a site of integration of human PRPCs 20 weeks post transplantation in a 43-week-old mutant xlpra2/RPGR dog (ID: Z720). A1–4 tdTomato-positive human PRPCs primarily differentiated into cone-arrestin (hARR3)-positive photoreceptors located into the host outer nuclear layer (ONL) and inner nuclear layer (INL). B All tdtomato-positive cells express human nuclear antigen (Ku80) confirming they are transplanted cells of human origin. Apparent tdTomato+/Ku80− signals reflect cytoplasmic extensions from donor cells overlying host nuclei, as confirmed by inspection of individual confocal z-sections. C1–2 Reconstitution of a continuous ZO1-positive external limiting membrane (ELM) after migration of human cone arrestin (hARR3)-positive PRPCs into the canine (host) ONL. D Transplanted PRPCs express human synaptophysin at their terminals and establish contacts with the host rod bipolar cells. (PKCα-positive). Scale bar = 20 µms
Taken together, these findings suggest that the persistence of at least three rows of nuclei in the host’s ONL may provide a more favorable environment for donor PRPCs, supporting their long-term survival and enabling a subset to differentiate into radially elongated photoreceptors. However, even at this less advanced stage of disease, the substantial early loss of donor cells following transplantation may account for the limited occurrence of successful integration events.
Three days after subretinal injection, transplanted hESC-PRPCs exhibit similar cell cluster profiles as cultured cellsTo investigate the factors contributing to the prominent early death of PRPCs after transplantation into the canine subretinal space, we conducted single-cell RNA sequencing (scRNAseq) on transplanted PRPCs. Approximately 10 million cells from stage 2 ROs (day 110–126) developed from hESC-derived WA09 CRX+/tdTomato line [21] were triturated into fragments and injected subretinally into systemically immunosuppressed healthy wild-type canine retinas. To evaluate whether mechanical dissociation itself compromised donor cell viability, we performed a LIVE/DEAD assay comparing RO cells before and after trituration. The results demonstrated no significant increase in cell death following dissociation (Supplementary Fig. 2), indicating that the observed cell stress and death in transplanted PRPCs cannot be attributed to preparative artifacts alone. Transplanted cell aggregates were visualized using optical coherence tomography (OCT) imaging 2 days PI (Fig. 3a), and harvested the following day (3 days PI) for scRNAseq analysis. This early time point was selected to maximize recovery of viable transplanted cells and enable identification of early markers associated with impending cell stress or death. For comparison, cells from the same batch of ROs were processed through dissociation but maintained in vitro for 3 days, allowing us to control for the effects of mechanical dissociation. To mitigate variability across batches, three independent transplantation experiments were performed.
Fig. 3
Single cell RNA sequencing (scRNAseq) analysis of donor PRPCs: a Near-Infrared cSLO image and single OCT b-scan showing the localization of subretinally-injected PRPCs (white asterisk) within the host retina two days post-transplantation. Green arrow indicates the location of OCT b-scan. b Schematic representation of the experimental workflow for isolating donor PRPCs and host retinal tissue for scRNAseq analysis. Created in BioRender. c UMAP plot illustrating major cell types identified in cultured and transplanted donor PRPCs. d Dot plot displaying key marker genes used for annotating cell clusters. e Proportions of each cell type identified in cultured versus transplanted PRPCs, derived from three independent experiments [mean (SD)]
To confirm that the analyzed cells originated exclusively from the transplanted human PRPCs and were not contaminated with host canine tissue, we aligned the same sequencing data to a combined human-canine reference genome and examined species-specific transcript expression. In one representative experiment, 11,022 cells were identified using the combined reference, of which 7,886 (71.5%) were classified as human, 6 (0.05%) as canine, and 3,121 (28.3%) as mixed, based on transcript content. Comparison with the human-only alignment revealed 7,336 total cells, with 6,426 barcodes (87.6%) overlapping between the two alignments. Similar distributions of species classification were observed in the two additional biological replicates. The presence of a small "mixed" population, including in samples cultured exclusively in vitro, suggests technical artifacts such as alignment to homologous sequences rather than true cross-species contamination. Importantly, all downstream analyses in this study were performed exclusively on cells aligned to the human genome alone, ensuring that only human donor cells were analyzed.
Additionally, we analyzed cells from the host canine retinas, comparing regions directly overlying the transplanted cells with regions where no PRPCs were present. The full study design is summarized in Fig. 3b. A total of 17,715 transplanted and 34,034 cultured cells were analyzed, leading to the identification and annotation of several retinal cell clusters, including rods, cones, horizontal cells and interneurons (HCIs), retinal ganglion cells (RGCs), retinal pigment epithelium cells (RPEs), Müller glia (MG), mitotic cells (MC), two distinct clusters of retinal progenitor cells (RPCs), and two clusters of neural progenitor cells (NPCs) (Fig. 3c). The cell clusters were assigned based on expression of groups of specific marker genes in the clusters (Fig. 3d). The percentage distribution of cells across these clusters showed no significant differences between the cultured organoids and transplanted cell microaggregates, however variability in cell numbers was noted across the three experiments (Fig. 3e). Notably, the percentage of RPE cells appeared elevated in the transplanted samples (14 vs. 3.6%). While this difference was not statistically significant, it may reflect subtle differences in RPE survival, enrichment, or early integration behavior in vivo and warrants further investigation. Overall, these findings align with our expectations, as no substantial differences in cell differentiation between cultured and transplanted cells were anticipated within the initial 3 days post-transplantation and batch-to-batch variability between retinal organoids has been noted earlier [19].
RNA velocity analysis reveals dynamic transcriptional trajectories in rods and cones during culture and following transplantationWhile the proportional distribution of cell types remained consistent between cultured and transplanted conditions at this early time point, we sought to investigate dynamic transcriptional changes within specific cell types during culture and transplantation. To this end, RNA velocity [29] analysis was performed to reconstruct transcriptional trajectories and assess maturation states in rod and cone photoreceptors. The analysis revealed dynamic transcriptional changes in rod and cone photoreceptors during differentiation in culture and following transplantation. ForceAtlas2-based [31] embedding of cone and rod populations (Fig. 4a–b, respectively) demonstrated a single trajectory of maturation under both cultured and transplanted conditions. However, RNA velocity vectors uncovered distinct dynamic differences between conditions and between the two photoreceptor types (Fig. 4c–d). In cones, RNA velocity vectors in the cultured condition pointed toward immature states, indicating a reduced commitment to maturation. Conversely, in the transplanted condition, vectors pointed toward maturation, with longer vectors observed in mature cells. This suggests enhanced transcriptional activity and a potential adaptive response to transplantation, allowing for greater functional integration in the host retina (Fig. 4c). In rods, transcriptional activity consistently pointed toward maturation under both conditions. Shorter RNA velocity vectors in mature cells in the cultured condition reflected reduced transcriptional activity, indicative of stabilization at terminal differentiation. In the transplanted condition, mature rods exhibited longer RNA velocity vectors, suggesting elevated transcriptional activity possibly due to adaptive responses to transplantation, efforts to integrate into the host retina, or activation of repair pathways (Fig. 4d).
Fig. 4
Continuous transcriptomic changes in human stem-cell derived cone and rod photoreceptors before and after transplantation: a–b ForceAtlas diagrams showing the integration of cultured and transplanted cone (a) and rod (b) photoreceptor subsets, illustrating full overlap of transcriptional states across conditions. c–d RNA velocity inferred trajectories for respectively cultured and transplanted cones (c) and rods (d), with arrows indicating the direction and velocity of transcriptional changes for each individual cell. e–h RNA velocity-based heatmaps highlighting variable genes along the transcriptional trajectories for cultured cones (e), transplanted cones (f), cultured rods (g) and transplanted rods (h), illustrating the progression of gene expression dynamics across maturation process
Heatmaps (Fig. 4e–h) further illustrated gene expression dynamics during photoreceptor maturation via unsupervised clustering of variable genes along transcriptional trajectories. In cultured cones, gene expression profiles showed a progression from genes associated with neuronal plasticity, transcriptional regulation, and stress responses (e.g., GAP43, CDKN1A, MEF2D, NFIA, USP48) to those involved in terminal differentiation, metabolism, and membrane dynamics (e.g., GUCA1B, GAD2, STMN2, GNAT2, DCX), indicative of increasing functional and synaptic maturity. Similarly, in rods, the gene expression profiles (e.g., RCVRN, GNGT1, RP1, IMPG1, PTPRZ1) demonstrated a transcriptional commitment to functional and synaptic maturation. In the transplanted condition, both rods and cones exhibited stress-related genes (GADD45A, CCL2, IFITM3, TXNIP etc.) alongside markers of cellular and synaptic maturation. This suggests that while transcriptional signatures of maturation persist post-transplantation, they are accompanied by robust stress responses likely reflecting adaptation to the transplantation process or the host retinal environment. Overall, these findings underscore the progression of photoreceptor maturation post-transplantation into the host retina.
Pathway analysis highlights oxidative and metabolic stress in PRPCs transplanted into normal canine retinasTo identify pathways contributing to increased cell death of transplanted PRPCs, we compared gene expression profiles between transplanted and in vitro cultured cells of each identified cell type. Differentially expressed genes were analyzed using Ingenuity Pathway Analysis (IPA) to identify enriched pathways specific to the transplanted cells. The top 30 enriched pathways identified through IPA, with Benjamini–Hochberg adjusted p-values < 0.05, are presented in Fig. 5a. Several pathways were consistently enriched across the different cell types.
Fig. 5
Pathway analysis reveals key biological processes impacted in cultured vs. transplanted PRPCs. a Ingenuity Pathway Analysis (IPA) identifies the top 30 significantly differentially regulated pathways in transplanted PRPCs, with Benjamini–Hochberg (B–H) adjusted p-values ≤ 0.05. b Heatmap of activation z-scores for 20 of the top 30 pathways highlights substantial metabolic dysregulation and oxidative stress in transplanted PRPCs. Gray boxes represent pathways or functions where IPA could not reliably predict the activation state. c IPA disease and functions analysis indicates an upregulation of cell death-related pathways in transplanted PRPCs, visualized as a heatmap of differentially regulated functions.
Most notably, 20 of these pathways (Fig. 5b) indicate that the majority of transplanted cells experience significant oxidative and metabolic stress. The top differentially regulated pathways, including Parkinson’s Signaling Pathway, Mitochondrial Dysfunction, and Sirtuin Signaling Pathway, are upregulated in all but one cell cluster. This widespread activation of stress-related pathways suggests that transplanted cells are mounting an adaptive metabolic response to the subretinal environment, engaging mechanisms linked to oxidative stress, mitochondrial maintenance, and cellular repair. In contrast, pathways essential for mitochondrial energy production, such as Formation of ATP by Chemiosmotic Coupling, Cristae Formation, Complex IV Assembly, Complex I Biogenesis, and Oxidative Phosphorylation, are markedly downregulated. Similarly, TP53 Regulation of Metabolic Genes and Respiratory Electron Transport pathways exhibit significant suppression, indicating compromised metabolic activity and diminished mitochondrial function. The observed downregulation of these pathways reflects a reduction in cellular energy production and metabolic efficiency, likely driven by the transition from a nutrient-rich culture environment to that of the subretinal space. Furthermore, the downregulation of Processing of Capped Intron-Containing Pre-mRNA suggests disruptions in nuclear processing critical for cellular function.
Although oxidative and metabolic stress pathways were enriched across all donor cell types, Müller glia and RPE cells exhibited the most significant transcriptional shifts, both in terms of pathway enrichment p-values and magnitude of activation/inhibition z-scores. This likely reflects their specialized roles in sensing and responding to metabolic stress and may signal the early initiation of a support response to stabilize the transplant environment.
Disease and function analysis in IPA (Fig. 5c) further supports these findings, showing increased activation of apoptotic and necrotic cell death pathways alongside decreased cell viability and survival. These trends are consistent across all cell types except the NPC2 cluster, where upregulation of energy production pathways (Fig. 5b) and greater resilience to the subretinal environment were observed, likely reflecting different energy demands and enhanced adaptability of this cell population.
Overall, these results suggest that the significant early loss of transplanted PRPCs is driven by metabolic reprogramming in response to a change in nutrient availability in the subretinal environment. This shift, coupled with oxidative stress and impaired mitochondrial function, likely contributes to the high rates of cell death observed shortly after transplantation.
Host retinal responses to PRPC transplantation are associated with metabolic reprogramming, immune activation, and translational adaptationTo characterize the host retinal response to transplanted cells, we analyzed retinal cells from normal canine hosts by comparing differential gene expression in cells directly overlying the transplanted PRPCs with cells collected from regions devoid of transplanted cells. Using expression of established cell-type markers, we identified all major retinal cell types, including rods, cones, rod bipolar cells, ON cone bipolar cells, OFF cone bipolar cells, amacrine cells, horizontal cells, retinal ganglion cells, Müller glia, astrocytes, and microglia (Fig. 6a–b). Pathway analysis revealed distinct changes in cellular processes among many host cell clusters, which were categorized into five functional groups (Fig. 6c): mitochondrial function and energy metabolism, protein translation and processing, amino acid regulation and metabolism, immune response and inflammation, and RNA processing.
Fig. 6
Single-cell RNA sequencing analysis of canine host neuroretina overlying transplanted cells compared to non-transplanted areas. a UMAP plot illustrating distinct cell clusters identified in the canine neuroretina overlying the graft, with each cluster corresponding to a specific retinal cell type. Similar clusters were also identified from neuroretina outside the transplanted region (not shown). b Dot plot illustrating expression levels of key marker genes used to annotate retinal cell types within the clusters. c Heatmap of top differentially regulated pathways, with Benjamini-Hochberg (B-H) adjusted p-values ≤ 0.05 identified by IPA, grouped into distinct biological categories. Gray boxes represent pathways or functions where IPA was unable to reliably predict the activation state
Within mitochondrial function and energy metabolism, Oxidative Phosphorylation, Respiratory Electron Transport, and Complex I Biogenesis were upregulated in host cells overlaying the transplanted PRPCs, suggesting an enhanced capacity for mitochondrial energy production. This is likely to reflect increased energy demands or compensatory mechanisms to counteract local stress. Paradoxically, Mitochondrial Dysfunction and Sirtuin Signaling Pathways were concurrently downregulated, which may indicate reduced activation of stress response and repair mechanisms essential for mitochondrial maintenance. This suggests that, despite an enhanced capacity for energy production, the ability to sustain mitochondrial health under stress may be compromised. Pathways related to protein translation and processing were broadly upregulated, including Nonsense-Mediated Decay (NMD), Eukaryotic Translation Initiation and Elongation, and Ribosomal Quality Control Signaling. These findings point to increased demands for protein synthesis and quality control, likely reflecting cellular efforts to adapt to stress conditions. Similarly, pathways involved in metabolic adaptation, such as Response of EIF2AK4 to Amino Acid Deficiency, EIF2 Signaling, and Selenoamino Acid Metabolism, were upregulated, highlighting nutrient stress and the need for tight regulation of amino acid availability. Immune response and inflammatory pathways were prominently activated, as evidenced by the upregulation of Neutrophil Degranulation and Neutrophil Extracellular Trap Signaling. These findings underscore a robust local inflammatory response at the transplant-host interface, likely driven by the presence of transplanted cells and not adequately suppressed by immunosuppression treatment. Additionally, RNA processing pathways, including Major Pathway of rRNA Processing and Processing of Capped Intron-Containing Pre-mRNA, were upregulated, suggesting enhanced transcriptional and translational activity to maintain cellular homeostasis under stress. Finally, the downregulation of Visual Phototransduction suggests localized dysfunction or reduced phototransduction activity in photoreceptors adjacent to the transplanted cells.
Interestingly, no significant pathway changes were detected in horizontal cells or retinal ganglion cells, suggesting that these cell types are not actively involved in responding to the transplanted PRPCs. Collectively, these findings reveal a complex interplay of metabolic reprogramming, immune activation, and translational adaptation in host retinal cells directly in contact with the PRPCs. The simultaneous upregulation of mitochondrial energy production pathways and downregulation of mitochondrial maintenance pathways underscores the metabolic strain at the transplant–host interface, while heightened inflammation and translational reprogramming reflect the cellular challenges of adapting to the altered microenvironment.
Immunohistochemical evidence confirms oxidative and cell stress responses in transplanted PRPCs and host photoreceptorsTo validate the transcriptomic findings from both transplanted PRPCs and the adjacent host retinal cells, we examined oxidative stress, mitochondrial integrity, and cell death markers using immunohistochemistry. This analysis enabled direct visualization of stress-related phenotypes and allowed us to determine whether the transcriptional changes observed in scRNAseq data were reflected at the protein level in situ. We stained for 4-hydroxynonenal (4-HNE, a marker of oxidative lipid peroxidation), ATP5A (ATP synthase F1 subunit alpha; a mitochondrial marker), cleaved caspase-3 (apoptosis marker), and HMGB1 (High Mobility Group Box 1, a nuclear protein that translocates to the cytoplasm during necrotic or inflammatory stress). For comparison, dissociated PRPCs from retinal organoids (ROs) were cultured in vitro for 3 days and processed similarly.
4-Hydroxynonenal staining was prominent in the host outer nuclear layer (ONL), particularly in photoreceptor inner segments and cell bodies, indicating oxidative stress in host photoreceptors overlying the transplanted PRPCs (Fig. 7A1). Transplanted PRPCs in the subretinal space also exhibited 4-HNE staining, though the signal was less intense and more variable (Fig. 7A2). In contrast, cultured PRPCs showed minimal 4-HNE signal (Fig. 7A3), suggesting that oxidative stress observed in vivo is driven by the host environment. Staining for ATP5A revealed strong and uniform mitochondrial expression in the host ONL (Fig. 7B1), consistent with intact mitochondrial structure and sustained energy production capacity in host photoreceptors. In contrast, ATP5A staining in the adjacent transplanted PRPCs (Fig. 7B2) appeared weaker and more heterogeneous, indicating early mitochondrial compromise. Cultured PRPCs displayed robust and uniform ATP5A staining (Fig. 7B3), similar to the host ONL, reflecting healthy mitochondrial function under in vitro conditions.
Cleaved caspase-3 staining (Fig. 7C1–2) revealed apoptosis in a subset of transplanted PRPCs, consistent with the IPA-predicted activation of apoptotic pathways. In contrast, neither host photoreceptors nor cultured PRPCs showed appreciable cleaved caspase-3 signal (Fig. 7C3), indicating that cell death at this early time point is largely restricted to donor cells. Similarly, HMGB1 exhibited cytoplasmic localization in transplanted PRPCs (Figs. 7D1–2), indicative of necrotic or inflammatory stress. Cultured PRPCs retained nuclear HMGB1 localization (Fig. 7D3), further confirming the in vivo specificity of the stress response.
Fig. 7
Immunohistochemical evidence of oxidative stress, mitochondrial dysfunction, and cell death in transplanted PRPCs. A1–A3 4-Hydroxynonenal (4-HNE) staining with (A1–A2) showing transplanted cells at 3 days post-injection and A3 showing cultured dissociated PRPCs. B1–B3 ATP5A staining in transplanted PRPCs (B1–B2) compared to cultured controls (B3). C1–C3 Cleaved caspase-3 staining apoptotic PRPCs in vivo C1–C2 versus absence of signal in cultured PRPCs (C3). D1–D3 Cytoplasmic HMGB1 staining in transplanted cells (D1–D2), consistent with necrotic or stressed states, while remaining nuclear in cultured PRPCs (D3). Scale bar = 20 µms
Together, these findings confirm that the subretinal environment induces early oxidative damage, mitochondrial dysfunction, and activation of both apoptotic and necrotic stress pathways in transplanted PRPCs. In contrast, host photoreceptors also exhibit oxidative stress but appear to avoid widespread cell death, suggesting that they may be better equipped to manage the metabolic and environmental stress induced by transplantation. These protein-level observations validate and reinforce the transcriptomic findings from both donor and host cell populations, underscoring the biological significance of early stress responses at the transplant interface.
Microglial infiltration and activation following PRPC transplantation into normal retina are primarily directed towards clearing apoptotic PRPCsMicroglia, the primary resident immune cells of the retina, demonstrated active infiltration into the outer retina and migration towards the PRPCs (Fig. 8A1–2). In contrast, microglia remained confined to the inner retinal space in regions devoid of PRPCs (Fig. 8B). Figure 8C1–3 show high magnification images of Iba1-positive microglia closely associated with tdTomato-positive PRPCs in the subretinal space. Several of these donor cells exhibit pyknotic nuclei (yellow arrows) indicative of apoptosis or necrosis. The spatial proximity and morphology suggest that microglia are actively engaging and engulfing dying PRPCs.
Fig. 8
Microglial response to donor human PRPCs in normal canine retinas 3 days post-xenotransplantation. Immunohistochemical analysis of microglial activation and migration in normal dogs (n = 2). Image shown is from dog LG5. A1–2 Host Iba1-positive microglia migrated into the host subretinal space and infiltrated the Td-tomato-positive donor PRPC mass. B In retinal regions distant from the transplantation site, microglia remain restricted to their native locations within the inner retina. C1-3 Higher magnification images showing Iba1-positive microglia apposed to tdTomato-positive donor cells with pyknotic nuclei (yellow arrows). D Ingenuity Pathway Analysis (IPA) of top enriched pathways in microglial, Müller glial, and astrocytic populations isolated from host retinas 3 days post-transplantation. Scale bar = 20 µms
To further investigate the role of microglia and other glial cells, such as astrocytes and Müller glia, in the host response, we analyzed pathways specifically upregulated in these cell populations. These pathways spanned five functional categories: immune response and inflammation, autophagy and cellular regulation, cell morphology and cytoskeletal dynamics, DNA synthesis and replication, and cell cycle and proliferation (Fig. 8d).
Immune response pathways, including Neutrophil Degranulation, Class I MHC-Mediated Antigen Processing and Presentation, and C-Type Lectin Receptors (CLRs), were significantly upregulated, indicating an active immune response likely driven by the recognition of transplanted cells as foreign or stressed. Additionally, the upregulation of the TNFR2 Non-Canonical NF-κB Pathway, NIK– > Noncanonical NF-κB Signaling, and T and B Cell Receptor Signaling pathways further suggests the engagement of both innate and adaptive immune mechanisms. These findings point to a coordinated immune activation within the retinal microenvironment, likely involving both resident immune cells and cellular signaling pathways typically associated with adaptive immunity, even in the absence of direct T and B cell infiltration.
Pathways related to cellular regulation and stress adaptation, such as Microautophagy Signaling and PTEN Regulation, were also upregulated, suggesting that glial cells were mobilizing regulatory mechanisms to maintain homeostasis in response to the transplantation-associated stress. Concurrently, upregulation of cytoskeletal and adhesion pathways, including Actin Cytoskeleton Signaling, RHOA Signaling, Integrin Signaling, and Signaling by Rho Family GTPases, indicated extensive cellular remodeling. These changes likely facilitate interactions with transplanted cells and contribute to reshaping the host microenvironment. Notably, upregulation of pathways involved in proliferation and DNA synthesis, such as Synthesis of DNA, DNA Replication Pre-Initiation, and mitotic phase-related pathways (Mitotic G2-G2/M Phases and Regulation of Mitotic Cell Cycle), suggests a robust gliotic response characterized by increased glial proliferation, potentially aimed at repairing or remodeling retinal architecture in the transplant region.
Together, these findings underscore a multifaceted host response to the transplanted PRPCs. The response is characterized by microglial activation, immune signaling, cytoskeletal remodeling, and glial proliferation, reflecting a coordinated effort to manage transplantation-induced stress and integrate the transplanted cells into the host retina. However, these processes also highlight potential challenges to graft survival and successful integration due to the complexity of the immune and cellular dynamics at the host-transplant interface.
Comments (0)