Tissue engineering approaches (including their combination with bioactive cells) represent promising treatment options to repair structural birth defects [20,21,22,23]. Yet, the application of these bioengineering technologies are invasive and only limited advancements have been made in these clinical scenarios [24, 25]. The stem cell therapy alternative, mainly involving cells isolated from gestational tissues, have been reported to improve animal survival and facilitate in utero tissue repair in genetically and mechanically induced spina bifida [8, 26,27,28]. Cell therapy poses several inherent risks and challenges, including issues with scalability, cell differentiation abilities and aging, bystander effect which reduces cell potency, number of cells reaching target sites, and therapeutic outcome [29].
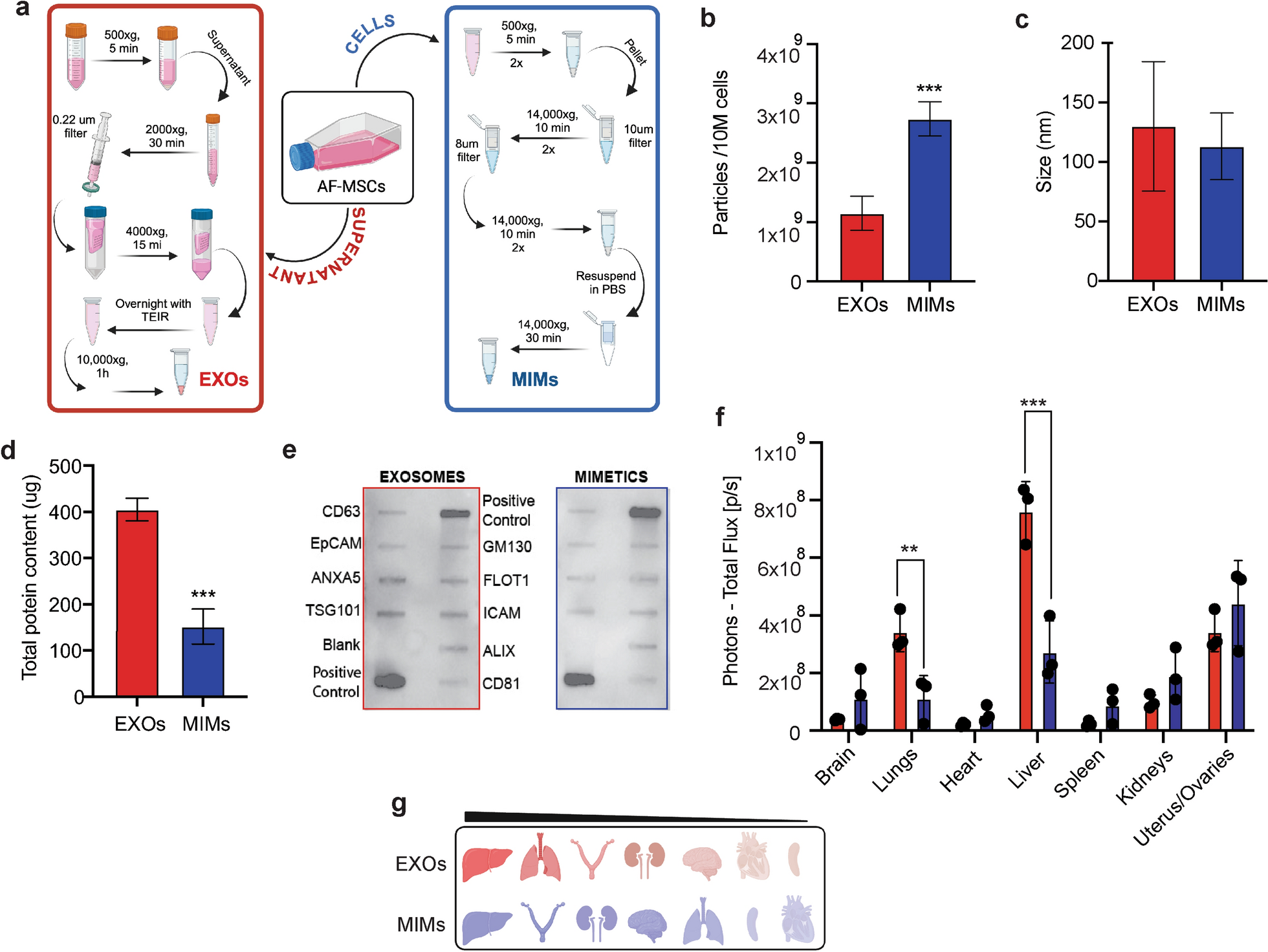
With this work, we present evidence suggesting that nanotherapeutics derived from AF-MSCs could serve as promising minimally invasive strategies for the prenatal repair of congenital anomalies. While EXOs from AF-MSCs have already been reported to retain parental cell molecular moieties and exert protective and regenerative potential [30, 31], we compared them to those associated to their mimetic counterparts produced from the same cells. MIMs were obtained through a recently established process [16], with the aim to potentially exploit them as alternative, therapeutic delivery systems. Our data suggest that both, EXOs and MIMs, could be used as scalable approaches to be tailored for individual clinical applications. Production of MIMs yields a 2.38-fold greater concentration than natural EXOs isolated from the same number of source cells (fresh AF-MSCs, 1 × 106). This trend is in line with previously acquired data, showing a 2–48 fold increase in the production of MIMs obtained from immune cells (IDEM) through the same process compared to natural EXOs [16]. To reduce variability and improve consistency across experiments, EXOs and MIMs were derived from the same batch, simultaneously exploiting the supernatant for isolation of EXOs and cells for MIMs. By utilizing a unified source for both particles, we minimized biological variability and ensured that any differences observed between the EXO and MIM populations were intrinsic to their biophysical properties, rather than being confounded by differences in cell batches or culture conditions. In terms of size, MIMs closely resemble their natural counterparts, with a diameter (113 ± 28 nm) which falls within the expected range for natural EXOs (130 ± 54 nm) [19, 32], although more heterogeneous when compared to IDEMs, and display a similar cohort of exosomal proteins. Recently, Sayyed et al. produced cell-derived nanovesicles from human adipose-derived-MSCs by cell extrusion with a mean diameter of 177.3 ± 2 nm and a yield of 1 × 1010 from 1 × 106 cells, surpassing the size produced in the current data, but showing a lower yield than was obtained in the present work [33]. On the other hand, Zhang et al. reported a mean diameter of mimetic nanovesicles of 126.9 ± 3.0 nm, with a yield of 16 × 109 particles from 1 × 106 cells, and a total protein concentration of 122.8 µg per batch [34]. Their yield and protein content were 20-fold higher than what they observed for natural EXOs. In our study, the yield of EXOs isolated from the same number of cells as MIMs was greater than the ones reported by these authors, although an overall similar trend in protein concentration was noted. MIM production from MSCs by Lu et al. generated EVs with a peak diameter mostly between 100–200 nm and a yield of approximately 1.6 × 106 [35]. It should be borne in mind that the diameter and composition of naturally secreted EXOs vary according to cell type and physiologic status, and environmental conditions, such as extracellular stimuli they are exposed to (including culture media and oxygen levels)[36]. Also, techniques used for EXO extraction, including variations in centrifugation protocols, type of rotor and g-force are aspects that play an important role in magnifying the yield, purity, protein content, and size of these EVs [37, 38]. Such variables should also be pondered in the production of MIMs and may explain differences encountered between the present results (in terms of yield, size and protein content) and data reported by others [33,34,35]. The feasibility of using frozen AF-MSCs to generate MIMs (F-MIMs) was evaluated to simplify the procedure by avoiding manipulation of fresh cells was also evaluated. Frozen cells produced a lower yield compared to fresh ones, but their diameter did not differ significantly, although the range of variation in the diameter of F-MIMs (min, 97.91, max: 119.01) was smaller than what was observed for MIMs (min: 85, max: 141). Cellular uptake evaluation demonstrated a reduction in the expression of GFP-positive macrophages over time, while a similar pattern of increased fibroblasts’ GFP expression was observed for MIMs and F-MIMs at 24 and 72 h, suggesting cryopreservation does not fully prevent the onset of apoptosis, impacting on cell recovery, which may explain the lower yield of F-MIMs and the differences observed when frozen cells were used [39].
The assessment of EV biodistribution is a pivotal stepping‐stone in the evaluation of their physiological significance, especially when it comes to the development of therapeutics [40], as it can determine the level of on‐target or off-target effects. Biodistribution is influenced by a variety of factors, including their cell origin, isolation techniques, and surface marker profiles [41]. To explore their biodistribution patterns and allow for a comparative analysis between naturally derived EXOs and engineered MIMs in terms of their organ-specific accumulation, fluorescently labeled particles were administered i.p. into female mice. While previous studies have reported the preferential accumulation of EVs in the lungs and liver following systemic and i.p. administration, although to different extents [42, 43], by looking into the female reproductive organs, our study reveals that EXOs and MIMs preferentially accumulate also in the uterus and ovaries. This unanticipated localization suggests AF-derived EXOs and MIMs have an inherent affinity or organotropism for reproductive tissues and may hold significant potential for targeted therapeutic interventions in the female reproductive system. The ability of naturally occurring EXOs to evade immune surveillance likely contributes to this biodistribution pattern [44]. Their small size, biocompatibility, and immune-modulatory surface proteins enable them to avoid rapid clearance by macrophages and other components of the innate immune system [14, 45]. This immune evasion mechanism not only enables EXOs to persist longer in circulation but also enhances their ability to accumulate in specific tissues where immune activity is naturally modulated. Based on this, our study suggests that MIMs may exhibit properties similar to those of EXOs; however, a more in-depth evaluation of their circulating levels at earlier time points is necessary to provide further insights.
Given the potential of EXOs and MIMs to specifically target uterine and ovarian tissues, we conducted a toxicology study using a well-established mouse model highly susceptible to drug-induced toxicity and NTDs, the SWV/Fnn strain [46]. EXOs or MIMs were administered i.p. every other day during a 15-day gestation period. Throughout the study, no significant changes were observed in the dams, as indicated by consistent maternal body weight and stable organ weights. Furthermore, the treatment did not impact the number of fetuses collected or result in a significant increase in fetal mortality or embryo resorption, suggesting that repeated administration of EXOs or MIMs during pregnancy is well tolerated in NTD-susceptible mouse model and supporting the safety of these vesicles for future therapeutic applications targeting reproductive tissues. The alterations produced by EXOs and MIMs in the hematological pattern of the SWV/Fnn females must be interpreted with caution, since these parameters may be influenced by variables such as age, gestation, liver and spleen functions, nutritional factors, among others [47, 48]. The MIMs reduced RBC and hemoglobin, increasing mean corpuscular volume, but these alterations did not lead to values outside the normal range for SWV/Fnn [49]. In addition, monocytes were significantly reduced by both EXOs and MIMs, but the greatest reduction was produced by EXOs when compared to MIMs. Most importantly, the reduction produced by MIMs (0.6 ± 0.2) kept monocyte counts above the norms for these animals (0.26 ± 0.17), whereas EXOs (0.17 ± 0.0) consistently reduced monocytes below the normal values, demonstrating that MIMs did impact innate immunity. To the best of our knowledge, this study is the first to comprehensively document the biodistribution of EVs in female mice, alongside the toxicological profile throughout gestation. In particular, the observed organotropism associated to EXOs and MIMs could be exploited for drug delivery specifically targeting uterine and ovarian tissues.
RNA encapsulation into EXOs represents a promising therapeutic strategy to various conditions, allowing for a more precise and ample control of protein expression than gene replacement therapy [50, 51]. In an effort to utilize both formulations for RNA therapeutic development, EXOs and MIMs were transfected with mRNA encoding GFP. Comparable values in terms of encapsulation rate were found between EXOs and MIMs (approximately 50%). This result contrasts with some reports in the literature, where mRNA loading into EXOs has been achieved with efficiencies as high as 90% [52] and highlights the influence of different loading strategies. Various methods, such as electroporation, passive diffusion, and specific transfection reagents, can significantly impact the loading outcome, leading to variability across studies [53]. Nonetheless, GFP-mRNA loaded within DiD-labeled MIMs and EXOs was efficiently delivered and expressed by two different cell lines, demonstrating the marked ability of MIMs to preserve mRNA functionality. Over time, we found that MIMs resulted in a greater percentage of GFP-positive macrophages and fibroblasts. In contrast, EXOs demonstrated a different temporal behavior within these cell populations. Specifically, the percentage of GFP-positive macrophages was initially higher during the first 24 h post-treatment with EXOs but decreased over time. However, in fibroblasts, the pattern of GFP expression induced by EXOs was similar to that observed with MIMs containing GFP mRNA. These findings suggest that the RNA cargo loading and delivery efficiency varies depending on the target cell type and function (i.e., varying capabilities for endocytosis and other uptake mechanisms) as well as on the type of nanoparticle used (i.e., composition, surface chemistry, size and shape [54]). Notably, MIMs were associated with a more prolonged expression of the delivered mRNA. The sustained and increasing expression of GFP mRNA delivered by MIMs indicates that this formulation may offer significant advantages over natural EXOs for mRNA encapsulation and delivery in therapeutic applications [16]. This differential uptake and sustained expression highlight the potential of MIMs as a more effective vehicle for mRNA-based therapies.
Next, we tested the plausibility of applying the technology of MIMs and EXOs as a potential therapeutic strategy for congenital malformations by using ex-vivo whole embryo culture. Our data showed that embryos explanted at E9.5 and cultured in a “soup” of DiD-labelled, mRNA-loaded EXOs or MIMs for 24 h, display the presence of a colocalization of signals at the level of the yolk sac where they expressed encapsulated GFP-mRNA. Importantly, no differences were found between the two formulations in support of embryo growth, yet only EXOs reached the embryo. As a semi-permeable barrier that allows for the exchange of nutrients, gases, and signaling molecules between the mother and fetus, this finding is not surprising. EVs have been reported as paracrine mediators of labor and delivery [55]. The ability of nanoparticles to cross the placenta is influenced by several recognized factors. Among these, lipid charge plays a significant role, with cationic nanoparticles demonstrating a higher likelihood of placental crossing compared to their anionic counterparts [56]. Additionally, the size of the nanoparticles (smaller particles exhibit greater placental uptake) and the gestational age at the time of exposure are crucial determinants [57]. Furthermore, the utilization of sensitive detection and quantitation methods is essential for understanding the extent and impact of nanoparticle accumulation in these regions. This includes assessing how exposure duration, clearance, and dosage affect placental transfer and accumulation.
While the accumulation of MIMs at the level of the yolk sac deserves a more detailed evaluation on the molecular mechanisms detaining them from crossing the placenta, data obtained here suggest their potential role as reconfigurable drug delivery tools to prevent the teratogenicity caused by maternal intake of drugs known to be toxic for the fetus, such as various anti-seizure medications which remain a hurdle in the treatment of pregnant women with seizure disorders [58,59,60], and other non-epileptic conditions [61]. In these instances, the chronic use of such substances is usually warranted to obtain adequate seizure control during pregnancy, raising serious concerns for pregnant women and those in childbearing age [62]. Malformations caused by these drugs are frequently severe and include NTDs, congenital cardiac and craniofacial defects [63]. On the other hand, since EXOs reach embryonic tissues and are primarily expressed in its cranial and ventral portions, they may be best suited for the prenatal repair of NTDs and other birth defects by loading these nanovesicles with cell-specific cargoes such as proteins, lipids, and nucleic acids [64]. This is not to say that despite the lack of direct contact with the embryo, MIMs may still play a role in the delivery of such molecules and be used in prenatal regenerative medicine through targeted delivery of genetic material to cells at the yolk sac by crosstalk and intercellular communication, due to its role in embryonic development [65, 66]. Further studies are warranted to best understand these mechanisms.



Comments (0)