
Remember me
We first quantified the selective protonation achieved with the proposed selective-deuteration method involving medium-switching from an unlabeled H2O medium to a labeled H2O medium with 15N-labeled ammonium chloride, [2H, 13C]-labeled glucose, and [2H, 13C, 15N]-labeled amino-acid mixture using solution NMR for GB1. Figure 3 shows a comparison of the 2D 1H-13C HSQC spectra of selectively deuterated [13C, 15N]-labeled GB1 produced with the [2H, 13C, 15N]-labeled amino-acid mixture (red) and fully protonated [13C, 15N]-labeled GB1 (black) in (a) the 1Hα/13Cα region and (b) the aliphatic side-chain region. The control spectrum in black serves as a reference (see Fig. S1 for assignments of the side-chain aliphatic region), to which deuteration-labeling efficiency was quantified for the GB1 sample produced by the medium-switching deuterium-labeling method. For the deuterated [13C, 15N]-labeled GB1 sample produced with the amino-acid mixture, except for attenuated signals of Lys, signals for Hα atoms were maintained for all the residues in the selectively deuterated sample as we hypothesized (Fig. 3a). Furthermore, in the aliphatic region, about half of the side-chain peaks observed in fully protonated [13C, 15N]-labeled GB1 were eliminated in the selectively deuterated sample (Fig. 3b).
Fig. 3 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Overlay of 1H-13C HSQC solution NMR spectra of fully protonated GB1 (black) and selectively deuterated GB1 produced by the medium-switching method with the addition of the unlabeled amino-acid mixture in the medium before switching the medium and with that of the deuterated amino-acid mixture after switching the medium (red) in (a) 1Hα/13Cα and (b) the side-chain aliphatic region. Both samples were dissolved in 50 mM phosphate buffer (pH 5.5), and these spectra were obtained with a 600 MHz solution NMR spectrometer equipped with a cryoprobe. The fully protonated and selectively deuterated GB1 protein concentrations were 83 and 96 µM, respectively. Contour levels were adjusted based on the protein concentration for each sample to compare signal intensities
For more quantitative analysis, the peak intensities for the HSQC spectrum of the selectively deuterated GB1 were compared with those of the fully protonated [13C, 15N]-labeled GB1 to determine the level of deuteration for the 1H-13C pairs of each amino-acid. Table 1 summarizes the peak intensities for each amino acid of the selectively deuterated [13C, 15N]-labeled GB1 normalized by the corresponding peak intensities of fully protonated [13C, 15N]-labeled GB1. For the 2D 1Hα/13Cα correlation data in Figs. 3a, and 48 well-resolved peaks in 56 amino-acid residues were analyzed. It should be noted that the peak intensities of the selectively deuterated sample used in the calculation were normalized by a scaling factor, which is a ratio of the concentration of the selectively deuterated sample to that of the fully protonated sample. Quantitative analysis by intensity comparison (excluding overlapped signals) showed that except for Asn, Ile, Lys, Thr, and α2 of Gly, all other amino acids showing resolved signals (see Table 1) indicate high levels of relative 1Hα signal intensities above ~ 90%, compared with the corresponding signals of the fully protonated GB1 sample. For Asn, Ile, Lys, Thr, and α2 of Gly, the relative 1Hα signal intensities were 77%, 72%, 27%, 61%, and 47%, respectively. On average, the relative 1Hα signal intensities for all the residues in the selectively deuterated protein were ~ 90%. It should be noted that for some amino acids, signal narrowing effects due to the side-chain deuteration were likely to introduce relative intensity higher than 100% (see Table 1).
Table 1 Relative peak intensities for the aliphatic region in a 2D 1H-13C HSQC solution NMR spectrum of GB1 prepared by the medium-switching from H2O medium containing the unlabeled amino-acid mixture to H2O medium containing the deuterated amino-acid mixtureIt is noteworthy that the method improved the protein yield (58 ± 3 mg/L culture), compared with uniformly isotope-labeled GB1 obtained without medium-switching and without the addition of the deuterated amino-acid mixture (39 ± 2 mg/L culture; see Fig. S3). It is likely that besides the addition of amino-acid mixture, switching the culture medium to a fresh one just before inducing the protein expression contributed to increasing the protein yield. Even for proteins with low expression levels, yield improvement can also be expected by applying this medium-switching method.
Notably, the intensities of the side-chain peaks were suppressed or decreased, compared with those of the corresponding peaks for the fully protonated [13C, 15N]-labeled GB1, as indicated in Table 1. The results show that the aliphatic side chains of Ile, Leu, Phe, Trp, Tyr, and Val were almost completely deuterated with 1H intensities of 0–16%. Ala, Asp, Asn, Glu, Gln, Lys, and Thr showed notably attenuated side-chain relative intensity levels of 0−43%, except for Asp, Glu, Gln, and Met 1Hβ. Asp, Glu, and Gln showed considerably high relative 1Hβ signal intensities at 74–79%, while Met 1Hβ showed a signal intensity of ~ 100%. A separate analysis based on the mass spectroscopy (m/z = 6779.7, Fig. 4b) indicated ca. 66% of the side-chain deuteration level, which is consistent with a high deuteration level indicated by the NMR analysis. It should be noted that a similar analysis for a fully protonated sample indicated m/z = 6574.7 (Fig. 4a). We confirmed that the protein expression in H2O medium with 15N-labeled ammonium chloride, [2H, 13C]-labeled glucose, and [2H, 13C, 15N]-labeled amino-acid mixture without the medium-switching reduces the side-chain deuteration to 54% (Fig. S2c). Thus, the medium switching is necessary to optimize the deuteration level.
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.MALDI-TOF-MS results for isotope-labeled GB1 samples prepared (b, c, d) with and (a, e) without solvent/medium-switching, and (b, c) with and (a, d, e) without the addition of the deuterated amino-acid mixture. *The deuteration rate of side chains calculated from MALDI-TOF-MS results assumes that HN and Hα are 100% protonated
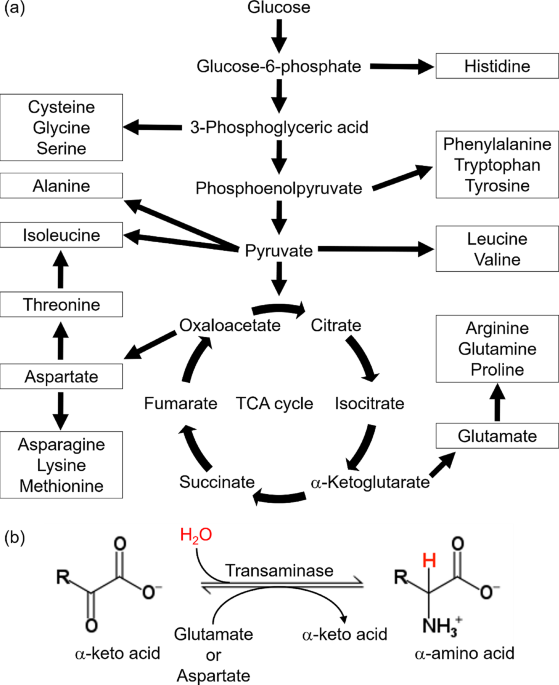
These results also suggest that the side-chain deuterium-labeling efficiency by this medium-switching method is amino-acid dependent. Amino acids such as Asp, Asn, Glu, and Gln with limited side-chain deuterium-labeling efficiency are known to be synthesized downstream in the TCA cycle (Maaheimo et al. 2001). The most compelling reasoning here is that the TCA cycle progressively produces back-protonated metabolic precursors such as oxaloacetate derived from deuterium-labeled glucose, incorporating protons from H2O into the side-chains of the amino acids that are synthesized downstream in the TCA cycle (Danmaliki et al. 2017). For example, the side-chain of the metabolic precursor corresponding to the β-position of Gln and Glu can be back-protonated when cis-aconitate is converted to D-isocitrate by aconitase via the incorporation of protons from the H2O solvent in the M9 media (Fig. 5).
Fig. 5 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.A reaction in the TCA cycle in which cis-aconitate is converted to D-isocitrate. In this reaction, protons derived from the solvent are incorporated into the skeleton from deuterated citrate
To examine the effects of the solvent before the induction, we also produced isotope-labeled GB1 protein using a solvent-switching protocol from isotope-labeled D2O medium with 15N-labeled ammonium chloride, [2H, 13C]-labeled glucose, and [2H, 13C, 15N]-labeled amino-acid mixture to labeled H2O medium containing the same reagents. As indicated above, this solvent-switching protocol was our original idea to improve the side-chain deuteration efficiency. With this protocol, we could obtain GB1 protein at a comparable side-chain deuteration level (67%), based on the mass spectroscopy data (6782.5, Fig. 4c), to that for the medium-switching method (66%; with the deuterated amino-acid mixture) using an unlabeled amino-acid mixture before medium switching and an isotope-labeled amino-acid mixture after medium switching. Figure 6 shows a comparison of the 2D 1H-13C HSQC spectra of side-chain deuterated [13C, 15N]-labeled GB1 produced using the solvent-switching method with the [2H, 13C, 15N]-labeled amino-acid mixture (blue) and fully protonated [13C, 15N]-labeled GB1 (black) in (a) the 1Hα/13Cα region and (b) the aliphatic side-chain region. For the deuterated [13C, 15N]-labeled GB1 sample, Hα atoms are maintained for all the residues. With the solvent-switching method, we observed stronger 1Hα signals for Lys, compared with those in Fig. 3a for the medium-switching method, for an unknown reason. In the aliphatic region, about half of the peaks observed in fully protonated [13C, 15N]-labeled GB1 are eliminated in the selectively deuterated sample (Fig. 6b). Overlaying the medium-switching and solvent-switching spectra, we can see that the signal patterns are quite similar (see Fig. S4). This solvent-switching method requires D2O and twice the amount of the isotope-labeled reagents, compared with the medium-switching method. We also produced the protein using twice the amount of [2H, 13C]-labeled glucose and [2H, 13C, 15N]-labeled amino-acid mixture to check the side-chain deuteration level by MALDI-TOF-MS (see Fig. S2d). The deuteration level was noticeably decreased (63%), compared with our proposed medium-switching (66%) and solvent-switching (67%) methods. Our proposed medium-switching method has achieved a higher side-chain deuteration level, while reducing the total cost for the isotope-labeled reagents and D2O.
Fig. 6 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Overlay of 1H-13C HSQC solution NMR spectra of fully protonated GB1 (black) and selectively deuterated GB1 produced by the solvent-switching method with the deuterated amino-acid mixture (blue) in (a) 1Hα/13Cα and (b) the side-chain aliphatic region. Both samples were dissolved in 50 mM phosphate buffer (pH 5.5), and these spectra were obtained with a 600 MHz solution NMR spectrometer equipped with a cryoprobe. The fully protonated and selectively deuterated GB1 protein concentrations were 83 and 92 µM, respectively. Contour levels are adjusted based on the protein concentration for each sample to compare signal intensities
Quantitative analysis by intensity comparison (excluding overlapped signals) showed that relative 1Hα signal intensities are ~ 78% on average, compared with those for the fully protonated GB1 sample. Thus, most of the 2Hα atoms are replaced by 1Hα in this protocol. Except for Ile, Lys, Thr, and α2 of Gly, for which relative 1Hα signal intensities to those of the fully protonated GB1 sample were respectively 61%, 36%, 49%, and 43%, all other amino acids showing resolved signals (see Table 2) indicate high levels of relative 1Hα signal intensities above 80%. In addition, the intensities of the side-chain peaks are suppressed or decreased, compared with those of the corresponding peaks for the fully protonated [13C, 15N]-labeled GB1. The results show that the aliphatic side chains of Ile, Leu, Phe, Trp, Tyr, and Val are almost completely deuterated (≤ 5%). While 1H side-chain signals were not fully suppressed, Ala, Asp, Asn, Glu, Gln, Lys, and Thr show reasonable attenuation for side-chain 1Hβ and 1Hγ signals at 0−63% and 0–34%, respectively, except for Met 1Hβ, which was barely replaced by 2H. Those results are comparable to those of the medium-switching protocol from H2O to H2O. While twice the amounts of isotope-labeled materials are needed, the yield (77 ± 13 mg/L culture) for this protocol was improved, compared with that for the medium-switching protocol (58 ± 3 mg/L). Thus, this method has its merits with mildly improved side-chain deuteration (see Table 2).
Table 2 Relative peak intensities for the aliphatic region in a 2D 1H-13C HSQC solution NMR spectrum of GB1 prepared by solvent-switching from D2O medium containing the deuterated amino-acid mixture to H2O medium containing the deuterated amino-acid mixtureTo confirm the effect of the deuterated amino-acid mixture in the H2O M9 medium, we also performed the corresponding experiments for deuterated [13C, 15N]-labeled GB1 produced by the solvent exchange method without [2H, 13C, 15N]-labeled amino-acid mixture in the medium. Again, Hα atoms are back-exchanged from 2H to 1H in most amino-acid types (Fig. 7a). However, the spectral analysis of the aliphatic side-chain region suggests a high level of protonation; almost all of the 1H/13C correlation peaks except for the β-position of tryptophan remain (Fig. 7b). Furthermore, quantitative analysis of the side-chain signals (Table 3) also showed that the side-chain 1Hβ signals exhibit high relative intensities at ~ 80% while other side-chain 1H species exhibit relative intensities at 50–87%, except for Met 1Hε (26%). Mass spectrometry results (6651.0; estimated side-chain deuteration 25%) in Fig. 4d also indicate a low deuteration level, which is consistent with the NMR analysis results. Thus, employing the deuterated amino-acid mixtures is mandatory to prepare target proteins with higher rates of deuteration at 65% or higher, as shown in Fig. 4b, c.
Fig. 7 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Overlay of 1H-13C HSQC solution NMR spectra of fully protonated GB1 (black) and selectively deuterated GB1 produced by the solvent-switching method without the deuterated amino-acid mixture (green) in (a) 1Hα/13Cα and (b) the aliphatic regions. Both samples were dissolved in 50 mM phosphate buffer (pH 5.5), and these spectra were obtained with a 600 MHz solution NMR spectrometer equipped with a cryoprobe. The fully protonated and selectively deuterated GB1 protein concentrations were 83 and 77 µM, respectively. Contour levels are adjusted based on the protein concentration for each sample to compare signal intensities
Table 3 Relative peak intensities for the aliphatic region in a 2D 1H-13 C HSQC solution NMR spectrum of GB1 that was produced by the solvent-switching method without the deuterated amino-acid mixtureIt should be noted that the observed protonation levels for different amino-acid types are consistent with those obtained with an inverse fractional deuteration method in a previous study by Medeiros-Silva et al. (Medeiros-Silva et al. 2016), in which they produced ubiquitin in a similar H2O medium (15N-labeled ammonium chloride and [2H, 13C]-labeled glucose) without any solvent/medium-switching. We prepared GB1 protein in an H2O medium containing 0.5 g/L 15N-labeled ammonium chloride and 2.0 g/L [2H, 13C]-labeled glucose as described in their work. The mass spectrometry analysis (m/z = 6650.1; Fig. 4e) indicates the side-chain-deuteration level at 24%, which is also substantially lower than that in the medium-switching method (66%). This deuteration level is almost the same (6651.0; estimated side-chain deuteration 24%) as that of GB1 prepared with the medium composition after solvent-switching of our sample without the deuterated amino-acid mixture (see Fig. S5 about the solution NMR results, Fig. S6 about SSNMR results, and Table S2 about line width comparison based on the SSNMR results). We concluded that the deuteration efficiency can be notably improved by the medium/solvent switching methods with the deuterated amino-acid mixture, compared with that of the fractional deuteration method (Medeiros-Silva etal. 2016).
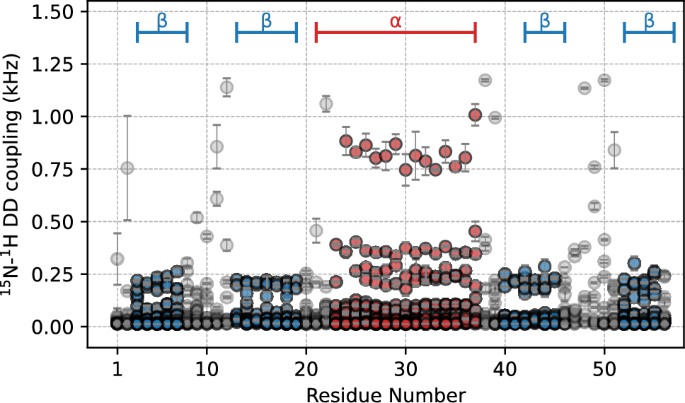
Resolution improvement by the solvent-switching methodNext, we examined improvements in 1H SSNMR resolution for samples prepared by selective deuteration with the medium- and solvent-switching methods. Figure 8 compares the 1H-detected 2D 1H/13C correlation SSNMR spectra of (a, c) selectively deuterated microcrystalline [13C, 15N]-labeled GB1 samples prepared with (a) medium-switching method with the unlabeled amino-acid mixture before switching the medium and [2H, 13C, 15N]-labeled amino-acid mixture after switching the medium (red) and (c) solvent-switching method with [2H, 13C, 15N]-labeled amino-acid mixture (blue) with a corresponding 2D SSNMR spectrum of (b) a fully protonated [13C, 15N]-labeled GB1 microcrystalline sample (black). The data were obtained with ultra-fast MAS at a spinning rate of 70 kHz. The spectra in Fig. 8(a–c) for the three samples showed 13 resolved peaks in 60 peaks amenable to quantitative analysis of 1H line widths (Table 4). The line widths of these 13 peaks were compared with the corresponding line widths of the side-chain-selectively deuterated sample to evaluate spectral resolution (see 1D slices in Fig. 8d–i). It was confirmed that, for almost all the resolved peaks, the 1H line widths of the selectively deuterated sample produced with the deuterated amino-acid mixture are narrower than those of the fully protonated sample, as shown in Fig. 8(d, e) (see Fig. S7 for full assignments of the 1Hα/13Cα region spectrum). The resolution improvement factors are in the range of (a) 0.92–1.94 and (c) 1.03–1.65 over the control data in (a); the average improvement factor is (a) 1.21 and (c) 1.22 (see Table 4). For Thr 51, the line width for the fully protonated sample is 166 Hz (Fig. 8f). In comparison, that of the solvent-switched deuterated sample is 156 Hz (Fig. 8j), resulting in an average resolution improvement by a factor of ~ 1.07. In some other residues, medium- and solvent-switching methods enable us to improve 1H line widths dramatically. For example, for Glu 27, the 1H line widths are (a) 131 Hz, (b) 251 Hz, and (c) 152 Hz (see also slices in (d, f, h)), resulting in significant resolution improvement by a factor of (a) 1.94 and (c) 1.65. Line width comparisons for other isolated peaks are described in Table 4 for the samples prepared with and without the [2H, 13C, 15N]-labeled amino-acid mixture, respectively. These results show that notable resolution enhancement can be achieved by the medium- and solvent-switching methods and supplementation with a suitable level of [2H, 13C, 15N]-labeled amino-acid mixture in the medium. For the inverse fractional deuterated sample, the resolution was mildly improved by a factor of 1.12 (data not shown). Thus, there is a notable advantage for the medium- and solvent-switching methods.
Fig. 8 The alternative text for this image may have been generated using AI.
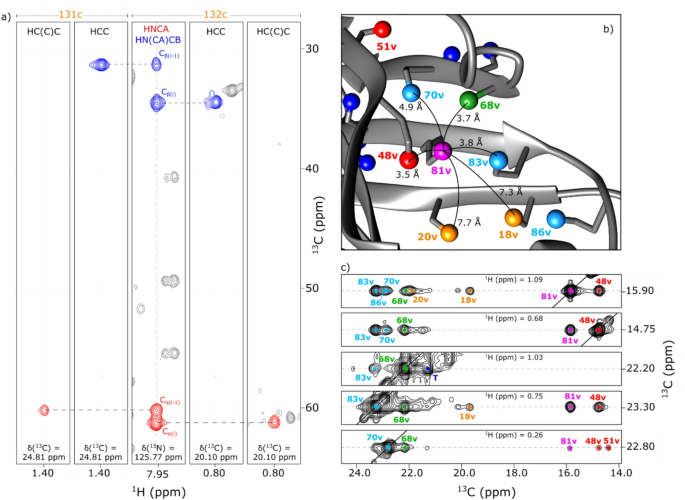
The alternative text for this image may have been generated using AI.Magnified 1Hα/13Cα regions in the 1H-detected 2D 1H/13C correlation SSNMR spectra of (a) a side-chain deuterated microcrystalline GB1 sample produced with the unlabeled before switching the medium and with the deuterated amino-acid mixture after switching the medium, (b) a fully protonated microcrystalline GB1 sample, and (c) a side-chain-deuterated microcrystalline GB1 sample produced by the solvent-switching method with the deuterated amino-acid mixture. (d−i) Comparison of 1D 1H slices selected at the positions of Glu 27 (13C chemical shift of 59.967 ppm; d, f, h) and Thr 51 (13C chemical shift of 62.571 ppm; e, g, i) for the same GB1 samples (d, e) selectively deuterated with the medium-switching method and addition of unlabeled amino-acid mixture before switching medium and the deuterated amino-acid mixture after switching the medium (f, g) fully protonated, and (h, i) selectively deuterated with the deuterated amino-acid mixture and the solvent-switching method. The spectra were obtained using a 900 MHz JEOL spectrometer with 70 kHz MAS. The 1D and 2D data were processed by NMRPipe/NMRDraw software
Table 4 1H line widths of side-chain-deuterated GB1 samples produced by the medium-switching method (with the addition of the unlabeled and deuterated amino-acid mixture in the medium before and after induction) and by the solvent-switching method (with the addition of the deuterated amino-acid mixture) and their resolution enhancement factors compared with a fully protonated GB1 sampleFor some amino acids, we observed higher suppression of side-chain 1H signals with a larger amount of amino-acid mixture (2.0 g/L; see Fig. S8 and S9). However, 1.0 g/L of the amino-acid mixture used for Fig. 3 was found to be sufficient to suppress side-chain 1H signals for most amino-acids (Fig. S10). Thus, we believe that the addition of 1.0 g/L of the amino-acid mixture is a recommended level for most applications of the medium-/solvent-switching method. Further optimization for a specific application is possible by changing the amino-acid composition of the amino-acid mixture or adding amino-acids with different isotope-labeling schemes.
Stereospecific assignment for Glycine residuesBased on solution NMR data, the ratio of the 1H signal intensity of α3 of glycine to that of α2 was estimated at ~ 2.0 with the medium-/solvent-switching methods as shown in Tables 1 and 2. The results are consistent with notable simplification of the SSNMR spectra in Gly region in Fig. 8(a, c). These results suggested that the α3 position in glycine was preferably protonated. Our method produced the target protein in H2O with a deuterated amino acid mixture and deuterated glucose. Loscha and Otting reported that the α3 position of protonated glycine was selectively deuterated by serine hydroxymethyltransferase by expressing the protein in D2O using protonated glycine (Loscha and Otting 2013). Glycine can be produced by serine hydroxymethyltransferase. Based on the reports by Movellan et al. (Movellan et al. 2019), we speculate that this reaction may involve transaminase and possibly other additional metabolic pathways. In any case, our method is likely to be useful for stereoselective Gly assignments.
Comments (0)