
Remember me
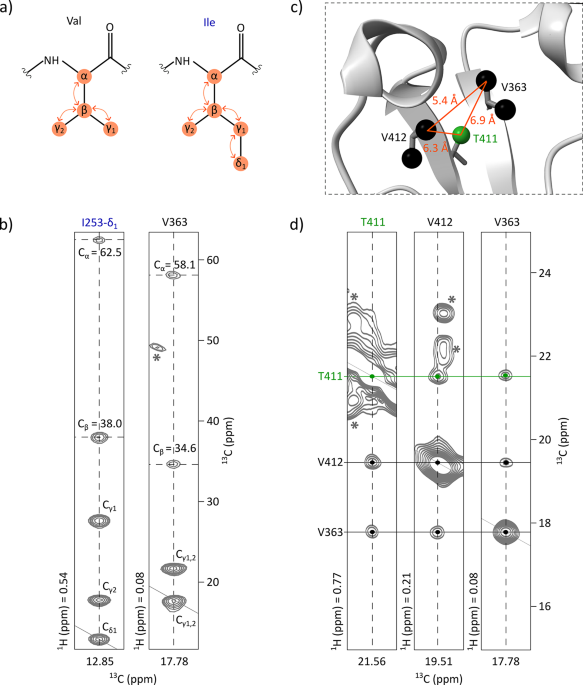
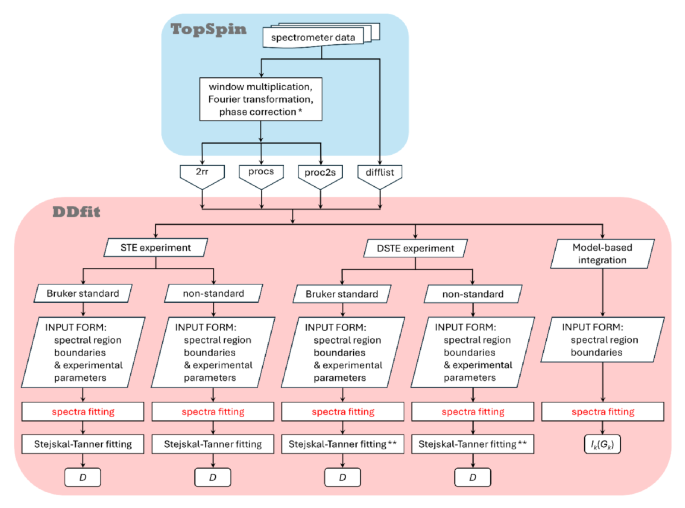
Two strategies were considered to assign the variable domains of ipilimumab Fab. We could study them either together as an ScFv fragment or separately using VL and VH domains. Working with the entire ScFv allows preservation of the interface between VL and VH as in the full Fab. However, the mass of ScFv is 29 kDa, and its assignment is complex without deuteration. Producing VL and VH individually gives smaller constructs that are easier to assign; however, the VH-VL interface is not preserved if the domains are produced separately, and some shifts can be observed upon transfer to the entire Fab. Addition of unlabelled VH to labelled VL could enable recovery of the interface between the two fragments. Ideally, obtaining both VL and VH domains soluble and concentrated is the most desirable option (see supplementary Fig. S5 for a flowchart diagram summarizing the overall strategy).
In our case, ipilimumab’s VL was easily produced, and its methyl group resonances were assigned. However, in our hands, the VH construct could not be obtained at concentrations enabling NMR sequential assignment. Protein precipitation occurred during sample concentration and upon buffer changes. Hence, production of the ScFv construct was also undertaken. In practice, although we have outlined both the advantages and limitations of each method, the final strategy was guided by the outcome of wet-lab production. With this divide and conquer method, the suitable size of the different constructs allowed us to produce them all in protonated buffer either uniformly 15N,13C-labelled or specifically methyl-labelled to assign each of these smaller Fab constructs.
Sequential assignments of both the light chain’s variable domain and the single-chain variable fragment of ipilimumab were performed using a combination of BEST and BEST-TROSY HNCA, HN(CA)CB, HNCO, HN(CA)CO, HN(CO)CA and HN(COCA)CB triple-resonance experiments (Favier and Brutscher 2019; Lescop et al. 2007) – see Table S3.
For the VL construct, 97% of 15N, 84% of Cα, 84% of C’, and 60% of the Cβ resonances were detected, excluding proline, tryptophan and cysteine residues. Indeed, the U-[1H, 15N, 13C] sample was produced in a cell-free system using a rich medium (hydrolyzed isogro®) that lacks free tryptophan and cysteine residues. Therefore, these latter amino acids were supplemented in the cell-free reaction, but only in their unlabelled form for cost reasons. Among the detected signals, 63% of 15N, 72% of Cα, 68% of C’, and 70% of Cβ could be assigned, tag included. Using hCCH-TOCSY experiments with mixing times of 10 ms and 20 ms, a transfer from the assigned backbone resonances to the methyl groups was undertaken. Using Cα and Cβ resonances, 71% of the methyl groups of the VL construct could be assigned, tag excluded. To complete the VL assignment, a 3D HCH NOESY experiment was acquired (see Table S3). The previously assigned methyl residues, together with the observed NOE cross-peaks, allowed the assignment of the remaining five threonine and six leucine methyl groups (Fig. 4, left).
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Assigned 2D 1H-13C SOFAST methyl TROSY spectra of the VL fragment of ipilimumab (left) and the ScFv of ipilimumab (right). VL and ScFv fragments were U-[1H, 15N, 13C] labelled. Each signal is annotated with the corresponding residue number. Alanines-β, isoleucines-δ1, valines-γ-pro-R, leucines-δ-pro-S, threonines-γ, and methionines-ε methyl signals are depicted in green, red, orange, light blue, dark blue, and purple, respectively. In bold are resonances belonging to the heavy chain and in regular the ones belonging to the light chain. Residues numbers are underlined for resonances of I-γ2, L-δ-pro-R, and V-γ-pro-S
For the ScFv construct, 92% of 15N, 86% of Cα, 88% of C’, and 59% of Cβ were detected, excluding proline, tryptophan and cysteine residues. Among the detected residues, 58% of 15N, 62% of Cα, 59% of C’, and 65% of Cβ could be assigned. Using an hCCH-TOCSY experiment together with the backbone assignment of ScFv and the already assigned methyl spectrum of the VL domain, 56 out of 74 methyl residues (76%) of ScFv were assigned, tag excluded, either by transfer from the backbone to the methyl groups or by transfer from the VL to the ScFv. All methyl groups belonging to the light chain of the ScFv were assigned, except alanine 35v and valine 3v pro-R. Some shifts were observed between the spectra of the ScFv and the VL (Fig. 1c, S2d, S3c, S4c), complicating the assignment transfer, but these shifts were expected for residues in very close proximity to the variable heavy chain of the ScFv. Therefore, we introduced some VH at natural abundance into our labelled VL samples and followed the shifts as we reproduced the environment found in the ScFv (Fig. 1d, S2e, S3d, S4d). This allowed us to assign alanine 35v in the ScFv construct.
To complete the assignment of methyl groups belonging to the variable heavy chain of the ScFv, the 0.22 mM sample of the U-[1H, 15N, 13C] ScFv ipilimumab construct was re-employed to acquire a 3D HCH NOESY experiment. Nine additional methyl residues (13%) were assigned using methyl NOEs. Two alanine residues in the variable heavy chain, 23v and 97v, initially remained unassigned. However, only alanine 97v is in close proximity to both a phenylalanine and a tryptophan, supporting the presence of a ring current effect and explaining the downfield shift of its 1H chemical shift. Based on this observation, both 23v and 97v could be confidently assigned and the percentage of assigned methyl groups was further improved to 92%, tag excluded (Fig. 4, right). Assigning methyl group signal of a 29 kDa protein without deuteration was a significant challenge. The high percentage of assignment obtained could be further improved by detecting long-range methyl-methyl NOEs (Kerfah et al. 2015; Ayala et al. 2020). This latter would, however, require the complex large-scale production of a deuterated and methyl labelled sample.
Methyl group assignment of ipilimumab’s FabBy transferring the assignment from the anti-LAMP1 Fab constant part and ipilimumab’s ScFv to ipilimumab’s Fab we could transfer 91% of valine, 93% of alanine, 100% of methionine, 100% of isoleucine, 91% of threonine, and 80% of leucine residues’ assignments (see supplementary Fig. S5 for a flowchart diagram summarizing the overall strategy). As assignments were not complete for the anti-LAMP1’s constant part and ipilimumab’s variable part, the percentage of ipilimumab’s Fab assignment is lower than the percentage of assignment transfer. Nonetheless, we reached 81% of ipilimumab’s Fab assignment, tag excluded, without having to produce it deuterated (Fig. 5).
Fig. 5 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Assigned 2D 1H-13C SOFAST methyl TROSY spectrum of a Fab fragment from the antibody Ipilimumab. The spectrum is a composite figure created by superimposition of four 2D 1-13 C SOFAST methyl TROSY spectra of Ipilimumab’s Fab labelled on Met-[13C1H3]ε and Ala-[13C1H3]β; Met-[13C1H3]ε, Ile-[13C1H3]δ1 and Thr-[13C1H3]γ; Met-[13C1H3]ε and Val-[13C1H3]pro−S&R and Met-[13C1H3]ε and Leu-[13C1H3]pro−S&R. Alanines, isoleucines, valines, leucines, threonines, and methionines resonances are depicted in green, red, orange, light blue, dark blue, and purple, respectively. Each assigned signal is annotated with the corresponding residue number. The asterisk indicates an impurity. In bold are resonances belonging to the heavy chain and in regular the ones belonging to the light chain. Annotated with the letter “c” are residues belonging to the constant part and with the letter “v” residues belonging to the variable part. Residues numbers are underlined for resonances of I-γ2, L-δ-pro-R, and V-γ-pro-S. “c*” indicate resonances of leucines-δ-pro-R and valines-γ-pro-S from constant part. (a) Full 2D spectrum. (b) Zoom from panel a. (c) Zoom from panel b
The limitations of this method lie in the fact that some residues in the ScFv construct have different surroundings compared to the same residues in the Fab fragment. This could potentially induce chemical shift perturbations for residues in the variable part located at the interface with the constant part, thereby complicating the assignment transfer from the isolated variable part to the entire Fab. However, this interface between the variable and the constant parts is relatively limited and contains very few methyl residues. The variable regions of both the light and heavy chains are connected to their respective constant parts with a loop constituted of approximately 6–8 residues. In our case, five methyl groups from the variable region are located less than 6 Å away from the constant domains, and six residues from the constant domains are within 6 Å of the variable region. These eleven residues account for only a small fraction of the residues targeted for assignment transfer. As previously discussed, the assignment of the six residues belonging to the constant part and close in space to the variable part (alanine 110c, valines 109c and 162c and threonines 163c and 171c from the light chain and alanine 119c from the heavy chain) was easily transferred from the already assigned anti-LAMP1 Fab to ipilimumab (Fig. 1b, S2a/b, S4a/b). This observation could be explained by the high percentage of identity between the two Fabs and very similar sequences in the variable region close to the constant domain (Fig. S6). On the contrary, transferring the assignment of the five residues belonging to the variable part and close in space to the constant part using the ScFv is slightly more complex as some shifts appeared due to chemical environmental changes between the ScFv and the entire Fab, but they represent only 3% of the total residue number.
Comments (0)