
Remember me
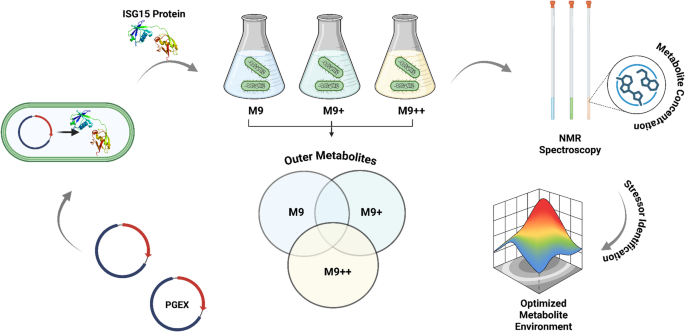
To investigate the influence of nutrients on E. coli biomass accumulation during recombinant protein expression, bacterial cultures which contain the PGEX-GST-ISG15 plasmid were grown in three defined media conditions: standard M9, M9+, and M9++. Final optical density at 595 nm (OD595) was recorded for each culture at the point of harvest, following IPTG induction and overnight incubation at 22 °C. All experiments were performed in biological replicates to ensure reproducibility. In the standard M9 minimal medium, the final OD595 values ranged from 1.75 to 1.99, with a mean of 1.87 ± 0.11(mean ± SD), reflecting restricted growth under nutrient-limited conditions. The limited availability of carbon, nitrogen, and cofactors in M9 medium is consistent with the observed low cell densities, which can constrain recombinant protein yield.
In contrast, cultures grown in M9 + medium, which contains defined supplements including additional phosphate buffering, glucose, and trace vitamins, demonstrated markedly enhanced growth. The OD595 values for M9 + ranged from 5.88 to 6.45, with a mean of 6.21 ± 0.24. This represents more than a threefold increase in cell density compared to M9, highlighting the critical role of nutrient availability in supporting bacterial biomass expansion during protein expression workflows. Interestingly, cultures grown in M9 + + medium, which contains further supplementation beyond M9+, reached a slightly lower mean OD595 of 5.42 ± 0.13. Although significantly higher than M9, the final cell densities in M9 + + were moderately lower than those observed in M9+. This subtle reduction may indicate the onset of metabolic feedback inhibition or osmotic stress resulting from excessive nutrient concentration, which can modulate growth kinetics under semi-defined conditions.
Overall, the data suggest that moderate enhancement of M9 medium (M9+) promotes optimal E. coli growth for recombinant protein production, while further enrichment (M9++) does not confer additional benefit and may slightly hinder maximal growth. These findings underscore the importance of empirically optimizing medium formulations to balance metabolic support with physiological tolerance during heterologous protein expression in E. coli.
Temporal metabolite profiling during recombinant protein expressionTo monitor dynamic metabolic changes during recombinant protein expression, samples were collected at three defined time points from each of the three biological replicates grown in M9, M9+, and M9 + + media. These time points corresponded to 20 min prior to IPTG induction (pre-induction), the midpoint of the expression period(mid-expression), and at the end of the expression phase (15 h post-induction). A total of 27 samples (3 media × 3 replicates × 3 time points) were analyzed by 1D 1H NMR spectroscopy (Fig. 2). After spectral acquisition, metabolite concentrations were quantified using Chenomx NMR Suite 12.00, employing DSS as an internal reference standard. This approach enabled absolute concentration determination for a panel of intracellular and extracellular metabolites. The NMR-based metabolic profiling revealed dynamic shifts in metabolite abundances across the expression timeline, which varied depending on the culture medium. Metabolomic analysis revealed distinct abundance profiles for key discriminatory metabolites among the three media conditions (Figs. 3 and 5). Acetate was found at the highest concentrations in M9, intermediate in M9+, and lowest in M9++, suggesting that overflow metabolism is most pronounced under minimal nutrient conditions and progressively reduced with increased supplementation. Formate displayed an opposite trend, with the highest levels in M9++, lowest in M9+, and intermediate in M9, indicating enhanced mixed-acid fermentation activity in the most enriched medium. Acetoin was also most abundant in M9++, present at intermediate levels in M9+, and lowest in M9, highlighting the activation of alternative fermentation pathways under nutrient-rich conditions. In contrast, citrate reached its highest concentration in M9++, was intermediate in M9, and lowest in M9+, suggesting increased flux through the TCA cycle or altered citrate utilization as medium complexity increased (Zhang et al. 2019). Collectively, these patterns underscore the profound impact of media composition on central carbon metabolism during recombinant protein expression in E. coli.
Fig. 3 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.The comparison of PLS-DA (top) and VIP scores (bottom) of M9, M9+, and M9 + + before induction (-), in induction (+), and further induction (++) without glucose among M9, M9+, and M9 + + individually
To further evaluate the significance of these metabolic changes, statistical analyses were performed using the MetaboAnalyst 6.0 platform. Comparative analyses focused on samples taken at the same time points across different media conditions, enabling the identification of metabolite variations specifically associated with media composition (Fig. 4). PLS-DA was employed to distinguish the key metabolites between media groups, and VIP scores were used to identify key metabolites contributing to the observed differences. This analysis revealed that certain metabolites—such as acetate, ethanol, and amino acids such as serine—played prominent roles in differentiating metabolic responses among M9, M9+, and M9 + + conditions. To specifically assess metabolic differences during active recombinant protein synthesis, a focused analysis was conducted on the mid-expression time point. At this stage, one sample from each of the three biological replicates was collected for each medium (M9, M9+, and M9++), yielding nine total samples for comparative evaluation. Multivariate statistical analysis using MetaboAnalyst revealed that metabolic profiles from M9 + and M9 + + conditions clustered closely, indicating similar metabolite compositions under these enriched media. In contrast, M9 samples showed a pronounced separation from both M9 + and M9++, suggesting a distinct metabolic state driven by minimal nutrient availability. Among the statistically important metabolites, formate, serine, acetoin, and ethanol were identified as the top contributors to the observed separation, as determined by VIP scores from PLS-DA. Acetoin concentrations were the highest in M9+, intermediate in M9++, and lowest in M9, possibly reflecting the impact of initial glucose supplementation and its downstream utilization. Conversely, formate levels were lowest in M9, moderate in M9+, and highest in M9++. Serine followed a pattern similar to acetoin and formate, with the highest levels in M9++, intermediate in M9+, and lowest in M9, supporting the hypothesis of increased overflow metabolism in nutrient-rich environments.
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.The comparison of PLS-DA (top) and VIP scores (bottom) of before induction (-), in induction (+), and further induction (++) among M9, M9+, and M9 + + without glucose
These results demonstrate that while M9 + and M9 + + induce relatively similar metabolic states during expression, standard M9 medium results in a markedly different metabolic response, particularly in central carbon and fermentation-related metabolites. Such differences may have implications for protein yield, solubility, and overall cellular health during recombinant production (Fig. 5).
Fig. 5 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Key metabolites’ main concentration values in induction (-), induction (+) and induction (++) stages for M9, M9 + and M9++
Late-stage expression reveals divergent amino acid and organic acid accumulation across mediaAt the final time point of recombinant expression (15 h post-induction), NMR-based metabolomic analysis revealed significant media-dependent variations in central metabolic intermediates. Among the most discriminatory metabolites identified by VIP scores from PLS-DA were ethanol, acetoin, formate, acetate, and valine, each displaying distinct distribution patterns across the three media conditions. Ethanol levels were highest in cultures grown in M9, intermediate in M9+, and lowest in M9++, suggesting greater fermentative activity in nutrient-limited conditions and a shift toward more respiratory metabolism as nutrient supplementation increased. Conversely, both acetoin and formate exhibited the opposite trend, with highest concentrations observed in M9++, moderate levels in M9+, and the lowest in M9, indicating enhanced mixed-acid and overflow fermentation activity in the more enriched environments. Similarly, valine followed the same distribution as acetoin and formate, with the highest accumulation in M9++, moderate levels in M9+, and minimal levels in M9. This pattern suggests that valine biosynthesis is promoted under nutrient-rich conditions, likely due to increased precursor availability and energy supply. Acetate, however, demonstrated a distinct inverse trend. Its concentration was highest in M9+, moderate in M9, and lowest in M9++, indicating that overflow metabolism may be more pronounced in M9 + despite moderate nutrient availability, potentially due to imbalances in carbon flux and redox state.
These observations collectively indicate that media composition profoundly alters the metabolic state of E. coli during the late phase of recombinant protein expression. While M9 conditions drive fermentative pathways marked by elevated ethanol production, enriched media such as M9 + and M9 + + shift metabolism toward enhanced biosynthesis and alternative fermentation end-products. Overall metabolites determined from induction and media conditions are demonstrated in Table 1. These metabolic reconfigurations likely reflect adaptive responses to differing nutrient landscapes and have implications for optimizing media formulations for improved protein yield and cellular health.
Table 1 Overall metabolite concentrations (µM) for M9, M9 + and M9 + + during Recombinant protein expression (induction (-) (before induction), induction (+) (mid of expression, and induction (++) (end of expression))Intra-medium metabolite fluctuations during the expression timelineIn addition to comparing media types at specific time points, temporal metabolic changes within each medium—pre-induction, mid-expression, and post-expression—were analyzed to uncover the dynamic progression of metabolic states during recombinant protein production.
In M9 minimal medium, the metabolites most strongly contributing to separation across time points were ethanol, betaine, valine, lactate, and putrescine. Among these, Ethanol, valine, lactate, and putrescine exhibited a consistent increase over time, suggesting progressive activation of amino acid biosynthesis and stress-associated pathways. In contrast, betaine concentrations steadily declined over time, possibly reflecting its consumption as an osmoprotectant under prolonged culture conditions. In M9 + medium, the most significantly variable metabolites were acetate, pyruvate, valine, and lactate. Acetate, lactate, valine, and pyruvate concentrations increased over time, consistent with elevated metabolic flux and amino acid synthesis during active expression (Vemuri et al. 2006; Wolfe 2005). Conversely, glycine decreased over the expression period, suggesting a shift away from fermentative pathways and potential glycine utilization for protein synthesis or one-carbon metabolism. In M9++, the key metabolites distinguishing expression stages were citrate, valine, leucine, adenine. Valine, adenine and leucine showed a progressive increase over time, aligning with enhanced biosynthetic capacity in this enriched medium. In contrast, both citrate ethanol displayed a steady decrease, potentially reflecting metabolic adaptation to stable osmotic and redox environments (Chae and Kim 2015). The results clearly demonstrate that media composition profoundly influences the metabolic state and growth dynamics of E. coli during recombinant protein expression. Specifically, standard M9 minimal medium, while advantageous for isotopic labeling due to its simplicity, significantly limits bacterial growth and induces a pronounced stress-related metabolic response. The elevated ethanol production observed in M9 indicates high fermentative activity, a common stress response due to nutrient scarcity and impaired respiration under minimal conditions. Although beneficial for specific labeling applications, such stress can negatively impact recombinant protein yield and solubility. In contrast, M9 + showed substantial improvement in bacterial growth and biomass accumulation. However, despite promoting high growth densities, M9 + displayed elevated acetate production, indicating pronounced overflow metabolism Enjalbert et al. 2015). Such overflow can reduce recombinant protein yield due to acidification of the medium and cellular toxicity, potentially limiting prolonged culture viability. Thus, while M9 + offers improved nutrient conditions compared to standard M9, its tendency toward overflow metabolism remains a critical limitation.
The M9 + + medium, designed with enhanced buffering and nutritional supplementation, provided intermediate growth levels between M9 and M9+, with significant metabolic benefits. Reduced ethanol and acetate concentrations suggest a more balanced metabolic state and lower stress, allowing a shift toward biosynthesis of valuable metabolites such as valine, formate, and acetoin. These metabolites potentially reflect enhanced cellular fitness, which is crucial for maintaining higher-quality protein expression. Despite these advantages, the slightly lower growth compared to M9 + suggests potential osmotic or nutrient imbalance stress from excessive supplementation in M9++. Optimization of glucose concentration, buffering agents, and precise vitamin supplementation in M9 + + may mitigate osmotic stress and further enhance bacterial performance. Additionally, adjustments to ammonium chloride and magnesium levels could optimize the balance between metabolic flux and cellular homeostasis, potentially improving overall recombinant protein yields.
Comments (0)