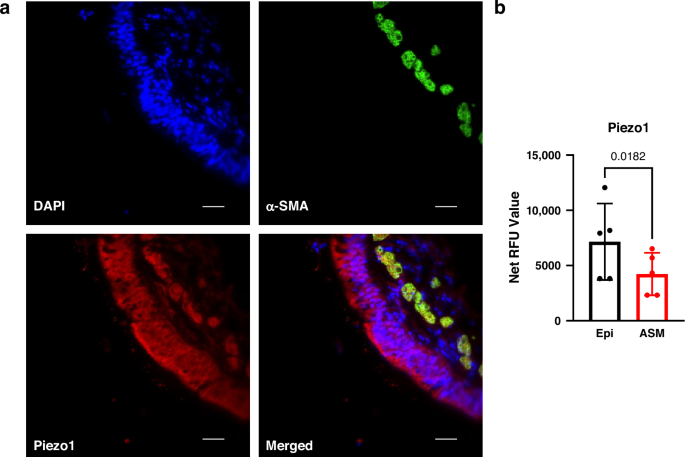
Recent evidence indicates that the central nervous system is closely associated with gut microbiota via the autonomic nervous system and humoral factors, including hormones, cytokines, and neurotransmitters. This bi-directional association, known as the brain–gut axis, has increasingly been implicated in neurological disorders, including epilepsy.5,20 Previous studies have reported an increased abundance of the Bacillota phylum (formerly Firmicutes) and decreased levels of Bacteroidota (formerly Bacteroidetes) and Actinomycetota (formerly Actinobacteria) in patients with epilepsy compared with healthy controls.18,21,22 However, prior investigations did not fully account for potential confounding factors.12,14,18,21,23 Lee et al. highlighted that clinical studies of gut microbiota in intractable epilepsy often produce inconsistent findings when considering confounders such as age, body mass index, and recent antibiotic exposure.12 Furthermore, as the gut microbiome is generally established around 3 years of age,24,25 studies including infants younger than this age may yield ambiguous results. Previous research on intractable epilepsy may also have been confounded by the direct effects of antiepileptic drugs on gut microbiota, similar to the antibiotic effect,26,27 as well as by the role of gut microbes in drug metabolism.28 More recently, several studies have analyzed the gut microbiota in patients with pediatric epilepsy prior to the initiation of treatment.9,10,11 Nevertheless, these studies are limited by their small cohort sizes, typically including fewer than 20 participants per study, which constrains the generalizability of their findings.
The present study had several advantages. We specifically targeted treatment-naïve gut microbiota in children with new-onset epilepsy aged ≥ 4 years and accounted for multiple confounding factors, including recent antimicrobial use, probiotic administration, and allergy-related conditions, which may potentially influence the intestinal microbiota. Furthermore, based on a priori power analysis, our sample size of treatment-naïve patients was sufficient to detect significant differences in the gut microbiota composition.
Using 16S rRNA gene sequencing, we compared the gut microbiota of 32 untreated children with epilepsy and 40 age-matched healthy controls, as well as longitudinal changes in eight patients for whom follow-up stool samples were available ~2–3 years after antiepileptic drug therapy.
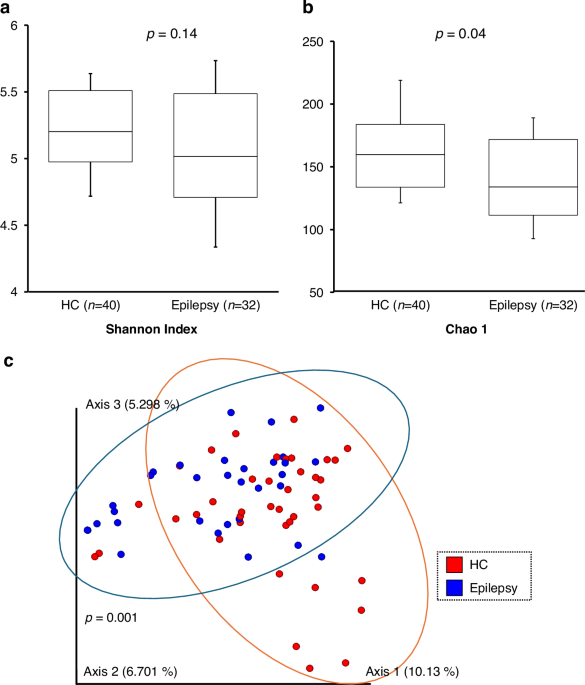
Our results revealed several key findings. First, children with treatment-naïve new-onset epilepsy exhibited significant dysbiosis, as reflected in both alpha and beta diversity metrics. Second, we expected that the beta diversity of the post-treatment epilepsy group (Epilepsy-post) would shift toward that of healthy controls following seizure control with antiepileptic drug therapy. Contrary to this expectation, the Epilepsy-post microbiota profile remained distinct from both the Epilepsy-pre and HC groups, suggesting that antiepileptic drugs themselves alter microbial composition. This effect is distinct from the well-documented impact of major dietary interventions, such as ketogenic diet therapy, which also profoundly shifts the microbiome.
Comparison of gut microbiota revealed an increase in Blautia and a decrease in Phocaeicola plebeius and Eisenbergiella, particularly in the epilepsy group. These differences may be significant because specific bacterial genera or species are beneficial for health, whereas others may contribute to disease pathogenesis, including epilepsy. For example, Blautia, the abundance of which was increased in children with epilepsy in this study, has been associated with metabolic and inflammatory disorders. However, it is also widely recognized as a producer of short-chain fatty acids, which have potential anti-inflammatory properties.29 This suggests a genus-level complexity where its precise role - beneficial or pathogenic - may depend on the specific species or environmental context. In contrast, Phocaeicola plebeius, enriched in the HC group, is generally considered a beneficial gut commensal with anti-inflammatory properties.30,31,32 The genus Eisenbergiella, although less well characterized, belongs to the Lachnospiraceae family, which produces butyric acid, a metabolite that strengthens intestinal barrier function and exhibits anti-inflammatory effects, suggesting that it may also contribute positively to gut health.
In this study, we used KEGG analysis to identify potential functional alterations associated with epilepsy. Rather than broad pathway names, we focused on specific bacterial genes to provide more precise insights. First, we found that the abundance of the pld gene, which maps to the glutamatergic synapse pathway in KEGG, was significantly reduced in the epilepsy group. Bannai et al. reported that calcium release from inositol triphosphate receptors and activation of metabotropic glutamate receptors and protein kinase C are required to stabilize the GABA type A receptor cluster.33 Second, the abundance of the tyrR gene was significantly lower in the epilepsy group. tyrR encodes a transcriptional regulator involved in tyrosine metabolism and isoquinoline alkaloid biosynthesis. While bacteria can metabolize tyrosine into various compounds, including tyramine and phenols, which may reduce the local tyrosine availability for host dopamine synthesis, the specific impact of gut microbial tyrosine metabolism on host neurophysiology is complex. The reduced abundance of tyrR-carrying bacteria suggests altered tyrosine regulation within the gut. Since tyrosine is a precursor for dopamine, such dysregulation could potentially influence downstream dopaminergic signaling. Previous studies indicate that dopamine plays a crucial role in modulating seizure activity34 and acts as a “remote control system” for seizure duration.35 Thus, the reduction in tyrR may be linked to altered dopamine modulation in epilepsy, although further metabolomic studies are needed to elucidate the precise metabolic consequences. Third, the bphB gene, associated with dioxin degradation, was also reduced. The direct link between bphB and epilepsy is unclear; however, this may reflect altered gut microbial capacity to metabolize environmental xenobiotics. These findings suggest that specific bacterial functional genes, rather than broad pathways, may contribute to the pathophysiology of epilepsy, although further multi-omics studies are needed to validate these predicted functions.
Interestingly, antiepileptic drugs significantly alter the taxonomic composition of the microbiota, as evidenced by distinct beta diversity clustering (Fig. 4c); additionally, they also induce a distinct functional reconfiguration (Fig. 5). We investigated whether the deviation in beta diversity observed in the post-treatment group could be explained by the type of antiepileptic drug administered. Among the six patients who deviated significantly from the HC and pre-treatment clusters, three received levetiracetam and three received valproic acid, suggesting that the deviation was not specific to the type of antiepileptic drug. Therefore, individual variability or other factors may play a larger role than drug type in shaping the gut microbiota in this small cohort.
Furthermore, our comprehensive functional analysis revealed significant alterations in several bacterial genes following treatment (Fig. 5). Genes involved in stress responses (dnaJ, jag, prdX) and metabolic processes (thiM, argD) were reduced, while those related to protein synthesis (tyrS, lspA, infC) were enriched. These findings suggest that antiepileptic drugs not only shift the taxonomic composition of the gut microbiota but also induce a distinct functional reconfiguration, potentially reflecting adaptation of the gut microbiome to the therapeutic environment. These functional adaptations might influence the metabolism or pharmacokinetics of antiepileptic drugs, potentially affecting the efficacy or tolerability profiles of specific medications such as levetiracetam or valproic acid.
This study has some limitations. First, detailed dietary intake data were unavailable, though probiotic intake was documented. However, dysbiosis in the pediatric patients with epilepsy confirmed in the present study appears to be independent of diet because of the following reason: previous studies demonstrating dysbiosis in epilepsy have focused on subjects with Western-style diets, while our study involved Japanese participants whose diets are presumed to be primarily Japanese. Second, the epilepsy cohort included various epilepsy types, which may have introduced heterogeneity. Gut dysbiosis may differ among specific epilepsy etiologies. However, our cohort size (n = 32) was not sufficient to perform subgroup analyses by epilepsy type. Nonetheless, the primary aim of this study was to identify a potential core dysbiosis signature in the shared state of treatment-naïve new-onset epilepsy before medication effects could confound the results. Future studies with larger sample sizes should investigate etiology-specific microbiota profiles. Third, this study relied on 16S rRNA gene sequencing, which has inherent limitations. We analyzed relative abundance rather than absolute bacterial load, so compositional changes may not reflect changes in absolute bacterial numbers. Whole metagenome sequencing would provide superior species-level resolution and direct functional gene identification. Finally, functional pathways were predicted using PICRUSt2. Although the shifts in several bacterial genes associated with epilepsy, such as bphB or tyrS identified by PICRUSt2 suggest a causative relationship between epilepsy and gut dysbiosis, it is just predictive analysis and therefore, a preliminary roadmap for future study. It is clearly needed to perform metabolomic or metatranscriptomic validation to add a layer of scientific rigor.
In conclusion, we compared gut microbiota composition between children with and without epilepsy. The findings suggest that dysbiosis—characterized by lower alpha diversity, distinct beta diversity, and enrichment of the genus Blautia, and significant reductions in Phocaeicola plebeius and Eisenbergiella—may contribute to the pathogenesis of epilepsy. Furthermore, comparison of pre- and post-treatment samples indicates that functional adaptation of the gut microbiota, potentially leading to changes in microbial metabolites, may be more relevant to seizure suppression than restoration of a specific “healthy” bacterial composition. Further studies, including metabolomic analyses, are needed to test this hypothesis. Future therapeutic strategies may involve correcting dysbiosis through prebiotic or probiotic supplementation or fecal microbiota transplantation from healthy donors.


Comments (0)