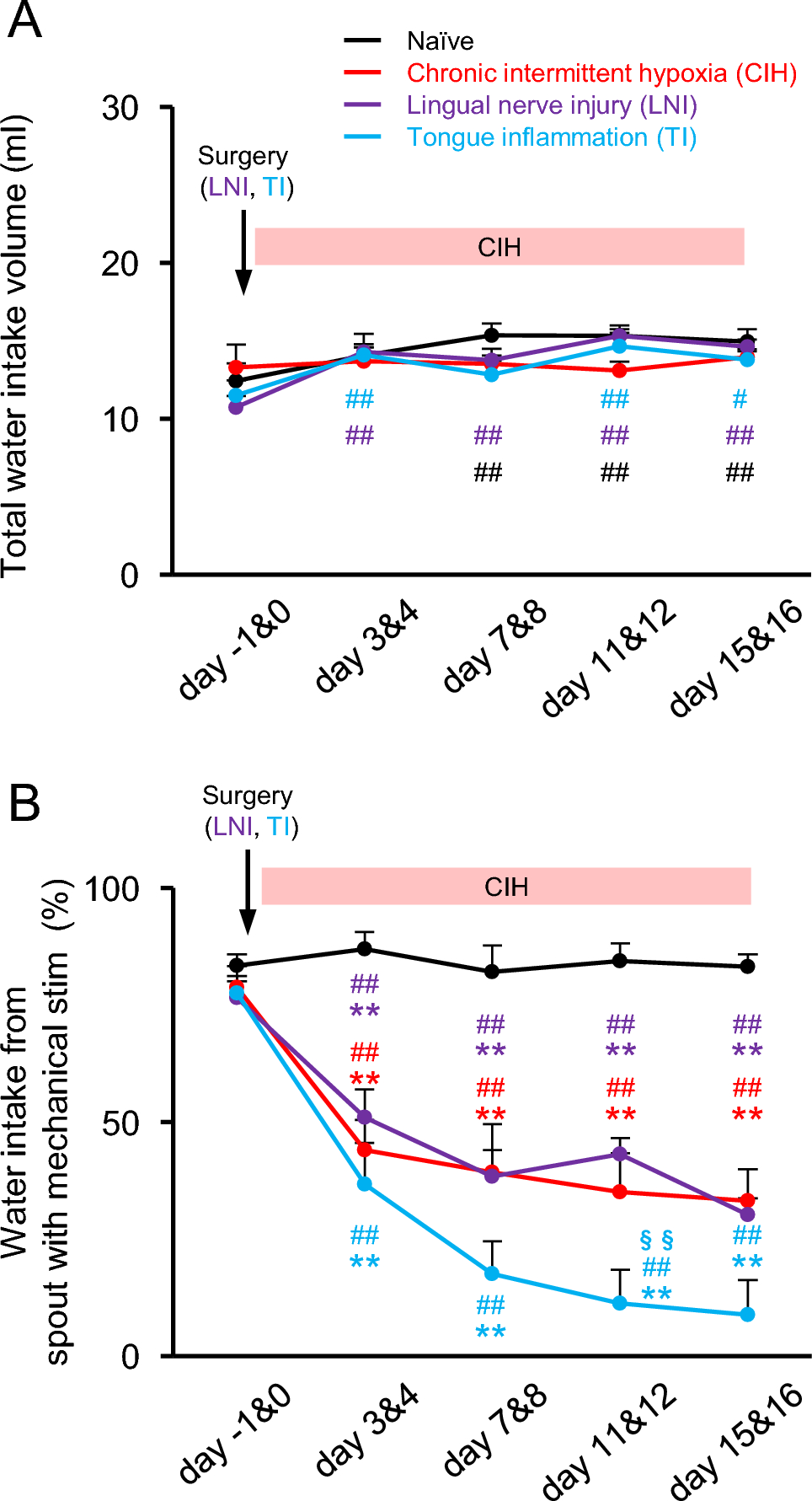
The present study demonstrated persistent mechanical allodynia of the tongue in rat models of CIH, LNI, and TI. This suggests that diverse pathological conditions affecting the orofacial region converge on common nociceptive signaling pathways. The molecular changes in the TG and Vc differed among these models, indicating that each condition engages distinct mechanisms of peripheral and central sensitization.
While this study provides substantial insights into pain-related mechanisms, the immunohistochemical analyses were conducted at a single time point and relied exclusively on marker expression to characterize neuronal and glial activation. Although this approach offers a clear and informative overview of molecular changes across the CIH, LNI, and TI models, it also underscores a critical avenue for future research: ascertaining whether these molecules actively contribute to the development or persistence of mechanical allodynia. Pharmacological or genetic interventions that selectively inhibit or modulate each molecule or receptor will be crucial for establishing their causal roles and confirming the functional significance of the observed changes. Furthermore, longitudinal analyses will elucidate the dynamic progression of these markers, including astrocytic and microglial responses in the Vc and phenotypic changes in TG neurons during the initiation and maintenance phases of pain [8, 14].
Initially, the rats exhibited a slight preference for spouts equipped with mechanical stimulation, which may indicate exploratory behavior or a preference for novelty in spout geometry. Notably, this baseline preference remained consistent and did not affect the interpretation of avoidance behaviors following CIH, LNI, or TI. Previous research has indicated that the thresholds for the escape reflex in response to mechanical pinch stimulation of the tongue under light isoflurane anesthesia return to baseline values, as observed in the control group, for the LNI and TI rat models after approximately 2 weeks [12, 13]. However, the current findings demonstrate that marked mechanical allodynia of the tongue persists for >2 weeks. The two-bottle choice drinking test conducted under free-moving conditions enables a reliable assessment of mechanical allodynia in the orofacial region, even in situations where central sensitization is minimal and peripheral sensitization is likely to be the primary contributor. Although prolonged mechanical allodynia provides a reasonable explanation for the sustained avoidance behavior observed in LNI and TI rats, no increase in cFos-IR cells was found in the Vc of LNI and TI rats. It is also possible that transient pain experienced during the initial phase led to a learned aversion to the spout with mechanical stimulation, which persisted even after the resolution of acute hypersensitivity.
In the present study, the optical fiber was positioned at the edge of the drinking spout; therefore, mechanical stimulation may have been applied not only to the tongue but also to the lips during water intake. Although the tongue is the primary structure that contacts the spout, we cannot exclude the possibility that avoidance behavior reflects mechanical allodynia involving both the tongue and perioral tissues. This potential confounding factor should be considered when interpreting the behavioral outcomes of the two-bottle choice test.
A major finding of this study was the enhanced expression of CGRP and HIF-1α in the TG of rats with CIH and TI. Increased CGRP expression is a hallmark of trigeminal nociceptor activation and is commonly observed in migraine and inflammatory, orofacial, and neuropathic pain [14, 26,27,28,29]. CGRP contributes to neurogenic inflammation, sensitization of peripheral terminals, and amplification of nociceptive neurotransmission [30]. Its upregulation suggests that hypoxic stress may directly sensitize trigeminal neurons [9]. The parallel elevation of HIF-1α supports this hypothesis. HIF-1α is a master transcription factor that is induced under hypoxic conditions. It regulates genes involved in pain processing, including those encoding pro-inflammatory cytokines and ion channels [31, 32]. Previous studies have demonstrated that hypoxia enhances pain sensitivity by increasing transient receptor potential vanilloid 1 activity via HIF-1α-dependent signaling in sensory neurons [16]. HIF-1α is involved in the increased expression of CGRP and activation of SGCs in the TG of a migraine model [29]. Therefore, the co-upregulation of HIF-1α, CGRP, and GFAP suggests that CIH triggers a sensitization pathway that enhances the excitability of nociceptors.
Our previous research demonstrated that CGRP expression in the TG increased following LNI; however, this elevation was confined to the early phase and returned to baseline levels by day 15. Phenotypic changes in CGRP-positive neurons were observed on day 3 [14]. Given that the present study collected tissue on day 16, it is probable that both CGRP expression and phenotypic changes had already been resolved, aligning with the current findings that show no upregulation of CGRP.
TI, but not LNI, increased HIF-1α expression in the TG. Hypoxia can induce inflammation, and inflamed lesions often become severely hypoxic due to immune cell infiltration and increased metabolic demand [33]. Hypoxia-mediated mechanisms may be an underappreciated component of inflammatory pain in the tongue. These results suggest that hypoxia-responsive gene networks may be common mediators of orofacial pain conditions involving systemic hypoxic stress, CIH, or localized inflammatory microenvironments.
Another key finding was the increased number of Piezo2-IR neurons and GFAP-positive SGCs in rats with CIH. Piezo2 is a mechanotransduction channel essential for tactile sensation [34], and previous studies have suggested its involvement in mechanical allodynia in neuropathic and inflammatory pain models [17, 35, 36]. Although the current study did not directly evaluate the function of Piezo2, its upregulation under CIH conditions suggests that intermittent hypoxia may influence mechanosensory transduction pathways and potentially reduce the mechanical threshold necessary to activate nociceptors in the tongue. Conversely, the expression of Piezo2-IR in TG neurons did not differ significantly between naïve rats and those subjected to LNI or TI in this study. Previous research has documented increased Piezo2 expression in trigeminal neuropathic pain models, such as infraorbital nerve ligation, with this increase becoming apparent approximately 7 days after nerve injury [36]. The lingual nerve crush model employed in this study typically induces less severe nerve injury than ligation models, potentially leading to distinct molecular responses. Furthermore, inflammatory responses in the TI model may be transient, which could account for the absence of sustained changes in Piezo2 expression at the examined time point.
SGC activation, as indicated by increased GFAP expression, is widely recognized as a contributor to peripheral sensitization in the sensory ganglia [9, 12, 14, 15, 37]. SGCs release adenosine triphosphate (ATP), cytokines, and chemokines, which enhance neuronal excitability through bidirectional communication [3, 20, 37]. The selective increase in SGC activation in rats with CIH suggests that glial-neuronal coupling may be one mechanism through which hypoxic stress enhances pain sensitivity and promotes central sensitization.
TRPV4 expression increased in LNI and TI rats but not in CIH rats. TRPV4 is a mechanosensitive and osmosensitive ion channel implicated in inflammatory and neuropathic pain in the trigeminal system [18, 38, 39]. Although its functional involvement was not examined in this study, its selective upregulation in the LNI and TI models suggests that tissue injury and inflammation may preferentially engage TRPV4-related pathways. In contrast, CIH may rely more heavily on Piezo2-mediated mechanosensation than on TRPV4. This divergence supports the notion that mechanical allodynia may arise through multiple molecular pathways, depending on the etiology.
cFos is an immediate early gene widely used as a marker of central sensitization and sustained nociceptive drive to spinal or trigeminal spinal subnucleus neurons [40, 41]. For example, the injection of capsaicin into the tongue of naïve rats resulted in a significant elevation of phosphorylated extracellular signal-regulated kinase (pERK) expression in the Vc [42]. Capsaicin application to the tongue reduces cFos expression following lingual nerve transection; however, it enhances cFos expression after infraorbital nerve transection [43]. This suggests that the pattern of immediate-early gene activation is contingent upon the type and location of nerve injury. Additionally, in TI rats (CFA injection on day 8), mechanical stimulation of the inflamed tongue enhances pERK expression [13], whereas in the CIH rats, low-dose capsaicin increases pERK, whereas high-dose capsaicin suppresses it [8]. Contrary to these studies, the present experiment did not involve any chemical or mechanical stimulation of the tongue at the time of brainstem sample collection, as the aim was to evaluate the molecular changes associated with persistent mechanical allodynia under unstimulated conditions. Given that cFos and pERK are immediate-early markers that predominantly reflect acute neuronal activation, the absence of additional noxious inputs likely accounts for the lack of increased cFos expression in the LNI and TI groups.
The absence of increased cFos expression in LNI and TI rat models at this late time point (day 16) suggests that central sensitization may have resolved or was not long-lasting. It is also possible that the intensity of peripheral input in the chronic phase of LNI and TI was insufficient to induce detectable cFos expression at the examined time point. In contrast, CIH appears to induce prolonged central hyperexcitability. Intermittent hypoxia has been reported to cause long-lasting plasticity in brainstem respiratory and cognitive-sensory processing pathways through oxidative stress, synaptic facilitation, and neuroinflammation [6, 7]. The current findings extend these observations to orofacial pain pathways and suggest that repeated hypoxic episodes, such as those occurring in OSA, may contribute to chronic alterations in trigeminal nociceptive circuits [8, 9].
These findings are clinically significant. Patients with OSA often report spontaneous burning sensations, soreness, or dysesthesia of the tongue and other oral structures [1]. OSA has also been associated with chronic pain disorders, including temporomandibular disorders [44] and headaches [45]. The present results provide mechanistic support for the hypothesis that CIH contributes to orofacial pain both peripherally (via CGRP, Piezo2, HIF-1α, and SGC activation) and centrally (via persistent Vc hyperactivity). This dual sensitization mechanism may explain the refractory nature of oral discomfort in patients with OSA.
Persistent pain following LNI appears to be driven primarily by peripheral mechanisms involving TRPV4-mediated mechanotransduction rather than sustained central changes. Although TI-induced inflammatory pain was severe, it did not result in central hyperexcitability by day 16. All three models were associated with long-lasting mechanical allodynia; however, only CIH was associated with prolonged central sensitization. This distinction may have therapeutic implications. Treatments targeting central mechanisms (e.g., gabapentinoids and central-acting neuromodulators) may be more effective for CIH-related tongue pain, whereas therapies aimed at peripheral inflammation or TRPV4/Piezo2 signaling may be more suitable for pain resulting from tongue injury or inflammation.
The present study showed that CIH and LNI, but not TI, induced extraterritorial mechanical hypersensitivity in rats. This finding indicates that conditions that cause robust central plasticity lead to widespread allodynia. CIH causes persistent activation of the trigeminal nucleus and increases Piezo2 and GFAP expression, which is consistent with hypoxia-driven neuroinflammation and central sensitization [46]. Neuropathic pain induces ectopic discharges and glial activation, which propagate central hyperexcitability [47]. In contrast, inflammatory tongue pain remains largely peripheral, consistent with evidence that transient inflammation does not sustain long-range central amplification [48]. These findings suggest that widespread sensitization emerges only when persistent central mechanisms are engaged.
In conclusion, peripheral sensitization, characterized by increased CGRP, HIF-1α, TRPV4, and Piezo2 expression and SGC activation, appears to contribute to persistent mechanical allodynia of the tongue in different pain models. However, CIH uniquely induces sustained central sensitization within the trigeminal brainstem, distinguishing OSA-related pain from neuropathic and inflammatory tongue pain. These findings suggest that distinct hypoxia‑responsive and mechanosensory pathways, in conjunction with neuron-glia interactions, may contribute to the development and persistence of chronic mechanical allodynia of the tongue.

Comments (0)