The objective of the present study was to investigate the effects of proprioceptive stimulation, by means of multiple muscle vibrations, on the perception of trunk movement and actual trunk movement in healthy individuals, in sitting and standing positions. Muscle vibration of the trunk induced (1) perceptions of trunk movement in sitting and standing positions and (2) small-amplitude trunk movements in sitting and standing positions, and (3) the amplitude of trunk movement was moderately associated with the vividness of movement perception.
All participants perceived a sensation of trunk movement during the application of muscle vibration. However, the vividness of the perception of movement was variable between participants, with individual median scores of perceptions ranging from 1.2 to 8.5 out of 10, in sitting or standing positions. Variability in vibration-induced perception is very frequent, both during single (Beaulieu et al. 2020) or multiple vibration stimulations (Tapin et al. 2021). One reason for this variability in the present study could be the inter-individual differences in anatomy, such as variations in the size of muscle and surrounding tissues (bones, soft tissues…) (Taylor et al. 2017), that may be more important at the trunk than on peripheral segments. There was no statistically significant difference between the vividness of perception of movement between sitting and standing positions or between different muscle vibration conditions in sitting or standing. The positions (sitting vs. standing) did not impact the postural requirements or state of the muscles, such as length or tone, sufficiently to alter perception, though these characteristics are known to facilitate or inhibit the perception of movement of the stimulated segment (Schofield et al. 2015; Taylor et al. 2017). This suggests that using proprioceptive stimulation to induce a perception of trunk movements for any application could be performed in either a sitting or standing position, without affecting the perception of movement.
Contrary to our hypothesis, participants perceived their trunk moving in the direction of the shortening of the stimulated muscles, e.g. trunk extension during the vibration of spinal muscles (SPIN condition), in most trials. These perceptions of movement are opposite to the perceived movement when muscle vibration is applied on the upper or lower limbs (Roll and Vedel 1982; Cordo et al. 1995; Ferrari et al. 2019; Beaulieu et al. 2020). This difference in perception between limb and trunk muscle vibration could be explained by the fact that in the vast majority of studies on perception, muscle vibration was applied on a relaxed, supported limb, with no postural activity required (Roll and Vedel 1982; Cordo et al. 1995; Ferrari et al. 2019; Beaulieu et al. 2020), contrary to the present experiment on movement perception at the trunk. In the present study, participants were asked to report the segmental displacement of their trunk during the vibration but not the sensation of the displacement of the whole body. The trunk was not restrained and thus required muscle activity to maintain balance. Therefore, the perception of movement may have been influenced by the postural activity necessary to keep a stable trunk posture. This postural activity likely included isometric contractions of the stimulated muscles that may have inhibited the feedback of the lengthening of the muscle elicited by MV (Ceyte et al. 2007). A similar situation is the vibration of the tonically active triceps surae in standing, which generated a backward body leaning that was perceived as such by the participants who generated a postural reaction that prevented a perceived fall forward (Eklund (1972). However, one can assume that the perception was mostly due to the actual postural response. Indeed, perceptions and postural reactions have been shown in opposite directions when appropriately evaluated, at least for cutaneous stimulation. Forefoot stimulation generated a perception of forward body leaning when the body movements were restrained (Roll et al. 2002), while the postural response was backward when the body movements were free (Kavounoudias et al. 1998).
Similarly, postural responses to ankle muscle vibrations are often interpreted as a response to counteract the perception associated with muscle vibration, i.e. the vibration of the soleus generates a perception of forward body leaning due to the perception of illusory lengthening of the soleus, which is counteracted by an actual posterior body leaning (Kavounoudias et al. 1998; Ceyte et al. 2007). Finally, the vividness of movement perception increased with the amplitude of trunk movement, as shown by a moderate correlation between these variables. Many studies showed that muscle vibration can elicit strong perception of movement without inducing large movement or any movement (Ferrari et al. 2019; Beaulieu et al. 2020; Le Franc et al. 2021). Similarly, at the individual level, some of the participants of the present study (e.g. participants 8 and 14) did not show large trunk movements but had a strong perception of trunk movement (see Tables S1, S2 and S3, in supplementary data). It is thus highly plausible that the perception of trunk movement in the direction of muscle shortening was not solely due to the actual trunk movement, but mostly to the sensory message generated by muscle vibration.
Another potential reason for the opposite perception direction at the trunk and limbs could be the use of co-vibration. Co-vibration of muscles of the trunk may be considered vibration of agonist but also antagonist muscles, depending on the considered movement. For example, right and left rectus abdominis are agonists in movements in the sagittal plane, but antagonists in the frontal plane. Thus the trunk responses could have been due to AVRs, as suggested by Calvin-Figuière et al. (1999). The results of the trunk kinematics support somewhat the direction of trunk perception of movement, with the trunk actually moving in the direction of the shortening of the stimulated muscles, and a moderate correlation between perception and movement amplitude. Therefore, the effect of co-vibration of antagonist muscles on the direction of the perceived movement requires further investigation.
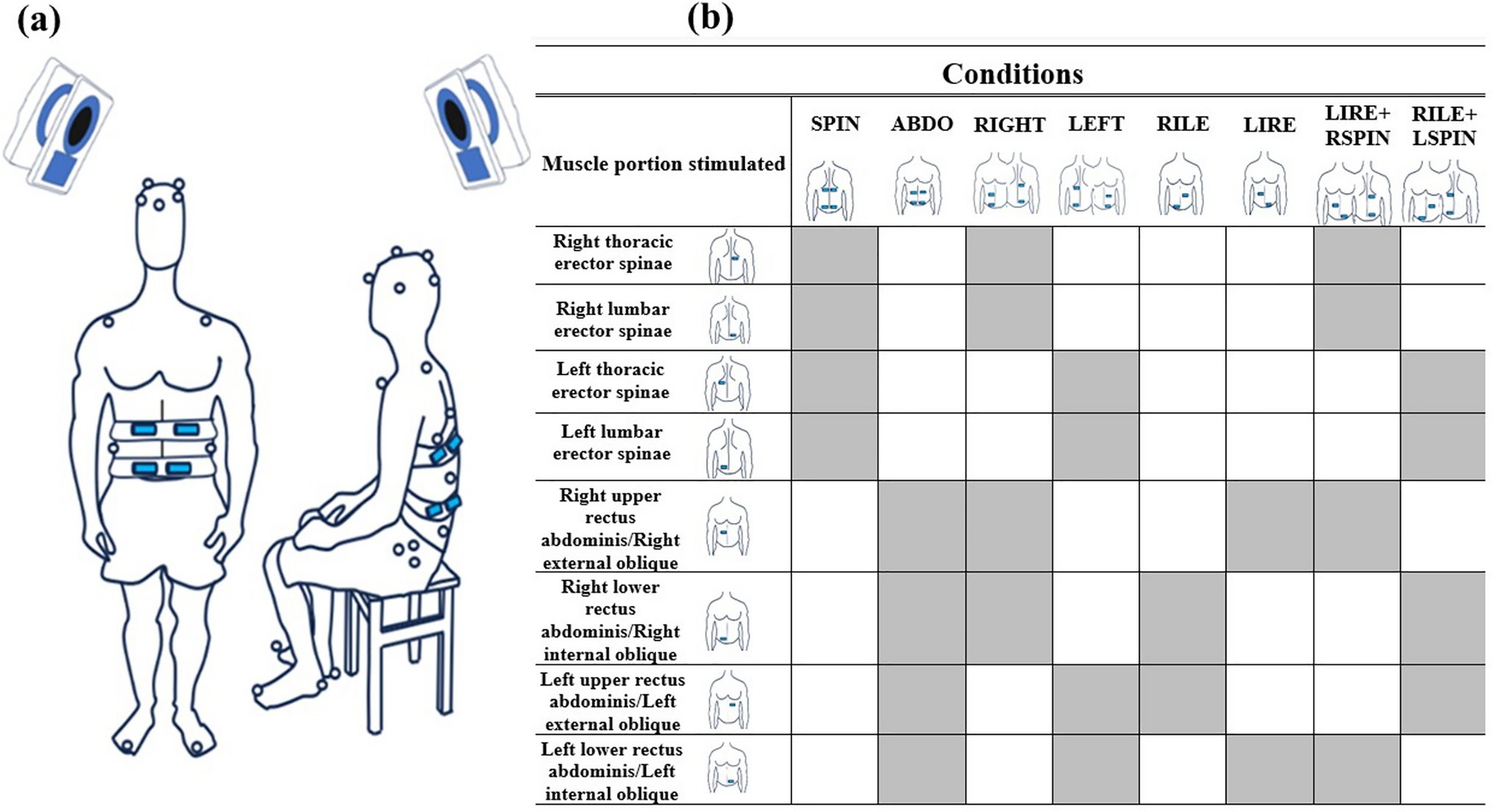
Additionally, perception of movements in unexpected planes, or no perception, is likely due to the vibration setup. First, the overlapping of abdominal muscles with various directions of action, such as the rectus abdominis and obliques, makes it impossible to specifically stimulate one of these groups to trigger a perception in only one specific direction. Similarly, the action of these muscles changes depending on which other ipsilateral or contralateral trunk muscles are co-activated, which likely alters perception at the trunk compared to the limbs (Bergenheim et al. 2000; Thyrion and Roll 2010). Finally, the number of vibrators used to generate the perception also possibly affected the perception, with more frequent unexpected plane or absence of perception in conditions targeting oblique muscles (RILE and LIRE conditions, i.e. right internal and left external obliques and left internal and right external obliques) that used only two vibrators, contrary to the four vibrators used in the other conditions (Taylor et al. 2017).
The application of multiple muscle vibrations also generated actual trunk movements. Proprioception is known to play a crucial role in the postural control of the trunk during both static conditions and dynamic tasks such as walking (Lekhel et al. 1997; Karnath et al. 2000; Bove et al. 2004). It is well documented that proprioceptive stimulation of the trunk and neck with a single vibrator can alter the spatial orientation reference system in dynamic tasks such as walking (Bove et al. 2004; Courtine et al. 2007). Applying vibrations to the neck while standing can also induce local movement in the sagittal and frontal planes in healthy individuals and individuals with neurological disorders (Karnath et al. 2000; Verrel et al. 2011). Courtine et al. (2007) showed that, during overground walking in healthy adults, muscle vibration applied unilaterally (left side) to various trunk muscles, including the lumbar and thoracic spinal muscles, trapezius, splenius, sternocleidomastoid, and abdominal muscles, induced global postural responses resulting in a curvilinear deviation of gait and a shift of the centre of pressure to the opposite side in standing. In the present study, the multiple muscle vibrations of the trunk elicited, for most trials, a shift in the direction of the stimulated side, which is opposite to the responses observed in the study just mentioned (Courtine et al. 2007). The nature of the task, i.e. sitting or standing vs. walking, in addition to the number of vibrated muscles, are thus factors that need to be further studied to understand how proprioceptive information is used to orient postural activity at the trunk.
Concerning the directions of movement, actual trunk movements were mostly in the plane related with the concentric action of the vibrated muscles. At first glance, this could appear to correspond to a TVR response of the trunk muscles, such as perceiving a backward trunk extension when erector spinae muscles were stimulated (SPIN conditions). Since muscle activity was not recorded in our study, it remains uncertain whether TVR occurred and could explain our findings.
For some other conditions, actual trunk movements occurred in unexpected planes. For example, the vibration of the right internal and left external obliques and left internal and right external obliques (RILE and LIRE) elicited the largest trunk movement in the sagittal plane. Such muscle vibration was expected to elicit larger trunk movement in the transverse plane. This response could be explained by the same reason as above, i.e. overlapping of the abdominal muscles (Taylor et al. 2017). Trunk rotation movement instead occurred mostly during the right and left trunk muscle vibration conditions (RIGHT and LEFT) rather than in conditions including the oblique muscles (RILE, LIRE, LIRE + RSPIN and RILE + LSPIN). Right and left trunk muscle vibration conditions elicited the largest trunk movement in the frontal plane. The rotation of the trunk mostly observed during RIGHT and LEFT conditions could be explained by the way that side bending of the trunk is generally associated with the rotation of the trunk (Levangie and Norkin 2001).
Limitations
Vibrators positioned on the abdominal muscles stimulated the upper portion of the rectus abdominis and the external oblique muscles at the same time (vibrators positioned at the top), the lower portion of the rectus abdominis and the internal oblique (vibrators positioned below). This likely increased the variability of the direction of the perception.
Another point was the difficulty of controlling the force of application of the vibrators. The elastic band used to maintain them was likely not always applied with the same tension. There was minimal tension to ensure the positioning of the vibrators in place but could have been higher for some participants. It may have affected the perception between participants, as this force can impact the perceptual responses (Ferrari et al. 2019).
It was also difficult to control the initial trunk position of participants. We could not restrain the trunk, since our objective was to allow trunk movements in all degrees of freedom. Small variations in initial position may have affected the length of each muscle and may thus have affected its response to vibration. However, the similarity in results between participants shows that the effect of these variations may be small. Finally, we used a non-validated numeric rating scale, whose psychometric properties are not known, to evaluate perception.


Comments (0)