
Remember me

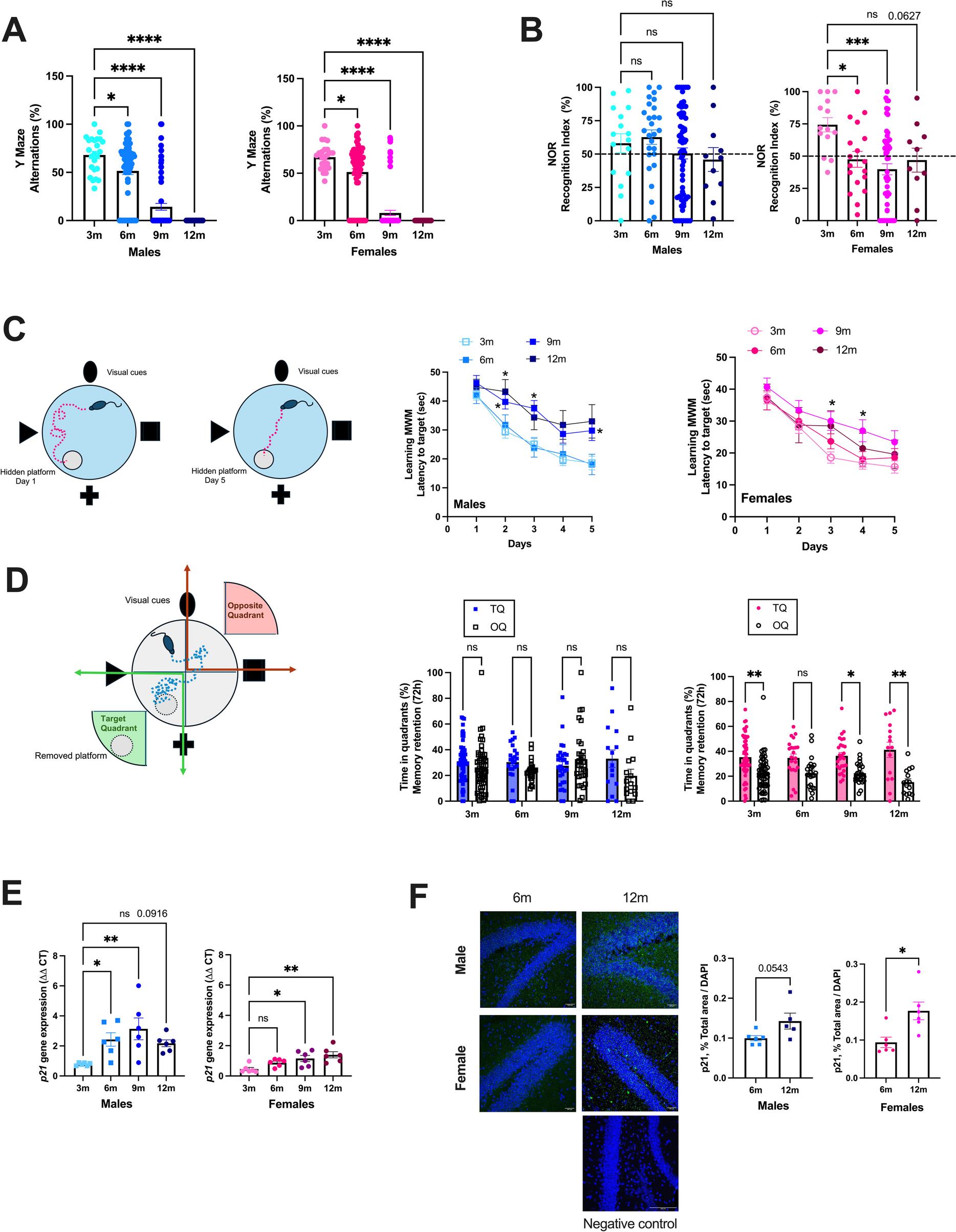


ATX mice are severely dyslipidemic [40, 42], are more frail and have a shorter life expectancy than their wild-type littermates [1]. Cognitive tests performed at different ages in naïve mice showed that spatial memory (Y Maze) decreased similarly with age in both male and female ATX mice (Fig. 1A). Spatial and recognition memory (NOR) tended to decrease with age in males and decreased in females as soon as 6 months of age (Fig. 1B). Learning memory assessed during the acquisition phase (MWM) evolved differently with age, according to the sex: in males, learning during the 5 successive days clearly slowed at 9- and 12-mo, whereas age had less effect in females (Fig. 1C). In addition, 12-mo female mice learned faster than males (day 2 in males 43 ± 4 vs. females 29 ± 6 s, n = 15, p < 0.05). Short-term (1 h) memory retention, assessed during probe trials in the MWM test, was unaffected by age or sex (data not shown). Male mice had no delayed (72 h) memory retention, which was only observed in females and not affected by age (Fig. 1D). This suggests that aging female ATX mice display a better delayed memory retention (probe trial), and to a lesser extent, a better learning memory (acquisition phase) than aging male mice. The cerebrovascular expression of the senescence marker p21 (cdkn1a), measured by RT-qPCR, tended to increase between the age of 3- to 12-mo by 180% in male (p = 0.092) and increased by 205% in female (p < 0.05) ATX mice (Fig. 1E). At both 6- and 9-mo, p21 expression was higher (p < 0.05) in males than in females. Immunofluorescence in the hippocampal region shows that the expression of p21 increased between the age of 6- to 12-mo by 44% in male (p = 0.054) and by 88% in female (p < 0.05) ATX mice (Fig. 1F).
Fig. 1
Learning impairment and increased p21 expression with atherosclerosis progression in ATX mice. (A) Spatial memory was assessed with the Y maze test in naïve 3 m (n = 24 males and n = 24 females), 6 m (n = 68 males and n = 64 females), 9 m (n = 70 males and n = 73 females) and 12 m (n = 18 males and n = 21 females) ATX mice. *: p < 0.05 vs. 3 m, one way-ANOVA and Dunnett’s multiple comparisons test. (B) Recognition memory was assessed with the novel object recognition (NOR) in naïve 3 m (n = 16 males and n = 14 females), 6 m (n = 27 males and n = 18 females), 9 m (n = 64 males and n = 54 females) and 12 m (n = 11 males and n = 10 females) ATX mice. *: p < 0.05 vs. 3 m, one way-ANOVA and Dunnett’s multiple comparisons test. (C) Learning memory was assessed with the Morris Water Maze (MWM) test during the acquisition phase over 5 consecutive days, in naïve 3 m (n = 55 males and n = 56 females), 6 m (n = 24 males and n = 24 females), 9 m (n = 30 males and n = 30 females) and 12 m (n = 15 males and n = 15 females) ATX mice. *: p < 0.05 vs. 3 m, Two-way ANOVA with repeated measures (Days X Age) and Dunnett’s multiple comparisons test. (D) Delayed memory retention assessed during the probe test 72 h after the removal of the platform in the MWM, in naïve 3 m (n = 55 males and n = 56 females), 6 m (n = 24 males and n = 24 females), 9 m (n = 30 males and n = 30 females) and 12 m (n = 15 males and n = 15 females) ATX mice. *: p < 0.05 vs. TQ, Two-way ANOVA with repeated measures (Quadrant x Age) and Sidak’s multiple comparisons test. Data are mean ± SEM of n mice. *: p < 0.05; **: p < 0.01; ***: p < 0.001; ****: p < 0.0001. (E) Gene expression level of the cerebrovascular senescence marker p21 was quantified by RT-qPCR in 3 m, 6 m, 9 m and 12 m male and female ATX mice (n = 6 mice per group). *: p < 0.05 vs. 3 m, one way-ANOVA and Dunnett’s multiple comparisons test. (F) Immunofluorescence of p21 in 6 m and 12 m ATX mice, in males and females (n = 6 images per age. Representative confocal images (20X), in addition to a negative control (40X), are shown; scale bar = 100 µm, for all images. *: p < 0.05 vs. 6 m, unpaired t-test)
Based on these data showing learning impairment and increased p21 expression with atherosclerosis progression, male and female ATX mice were treated with the senolytic ABT-263 (Navitoclax) using 2 time frames: a preventive treatment in young mice from 3- to 6-mo, or a curative treatment in middle-aged mice from 9- to 12-mo was applied, to respectively prevent or delay VCI in male and female ATX mice.
Treatment with ABT-263 was well toleratedAs expected, body weight was smaller in female than in male mice, but it was not affected by ABT-263 in both preventive and curative treatments (Figure S1). Globally, the preventive treatment with ABT-263 (from 3- to 6-mo) had little effect on blood markers of liver or renal function and did not alter the lipid profile or glucose levels, neither in male nor in female mice (Table S3). Only in females, ABT-263 lowered alkaline phosphatase. When compared to male-placebo mice, female-placebo mice had lower TG, but higher urea and higher alkaline phosphatase (Table S3). Late treatment with ABT-263 (from 9- to 12-mo) lowered cholesterol and TG only in female mice; placebo-female mice had higher glucose, cholesterol, TG and phosphatase alkaline (Table S3). These data suggest that, overall, the chronic intermittent treatments with ABT-263 were well tolerated by the mice.
ABT-263 improves cognition in male ATX mice onlySpatial working memory and exploratory behaviour assessed with the Y maze test were not affected by ABT-263 in young male and female mice (Figure S2A). Exploratory behavior decreases strongly with age, and at 12-mo, 100% of the mice failed the Y maze test in all groups (Figure S2A); indeed, none of the 12-mo mice, in either group, entered at least 8 times in the arms of the maze (Figure S2A).
Recognition memory measured with the NOR test was first validated in young, untreated, 6-mo male ATX mice (n = 23, Figure S2B): according to the inclusion criteria of the test, the time spent on both objects was higher than 20 s; during the training trial, the time spent on both objects was similar (p = 0.708) while during the test trial, the time spent on the novel object was significantly higher (p = 0.008), demonstrating that mice discriminate the novel from the familiar object and that the test was functional (Figure S2B). In these young, untreated male ATX mice, the recognition index was higher than 50% − the limit for a discrimination by chance − and considered satisfactory (64.9 ± 4.6%; n = 23). The preventive treatment with ABT-263 did not affect recognition memory in young mice of both sexes (Figure S2C). Middle-aged mice lacked curiosity for the novel object and ABT-263 did not improve recognition memory, as illustrated by a discrimination index close to or lower than 50% in all 12-mo groups (Figure S2C). The success rate for NOR in middle-aged mice was poor, in both males (55%) and females (30%). There was, however, no motor dysfunctions since neither total distance traveled, nor velocity of movement were affected by treatment or sex (Figure S2D, E).
We also used the MWM test, a more complex behavioral test performed over 11 days, targeting learning and spatial memory. Because mice must escape from the water, it works even in middle-aged mice lacking exploratory curiosity. Over the 5 days of the learning curve (D1-D5), mice learned to locate the immersed hidden platform and the latency time to reach the target decreased progressively. In young male mice (6-mo), similar learning curves were observed for placebo or ABT-263-treated mice (Fig. 2A). In 6-mo placebo male mice the learning process, performed in the same mice at 3-mo (Fig. 1C), did not reach statistical significance (6-mo placebo D5: 22 ± 5 s vs. D1: 33 ± 5 s, p > 0.05), but ABT-263 improved learning in male mice as they successfully located the platform on D4 (6-mo ABT-263 D4: 15 ± 4 s vs. D1: 25 ± 3 s, p = 0.0162) (Fig. 2A). In females, placebo- and ABT-263-treated mice exhibited similar learning pathways, they both located the platform on D5 (Fig. 2B). This suggests that ABT-263 was slightly more beneficial in young males, but ineffective in females.
Fig. 2
Impact of ABT-263 treatment on cognition in the MWM (A) Latency to find the hidden platform from day 1 to day 5 in the acquisition phase (learning memory) of the Morris water maze (MWM) test of male (M, A) and female (F, B) mice after treatment with placebo (P) or ABT-263 (ABT) at 6-month-old (6 m, A and B) and 12-month-old (12 m, C and D). Each value is a mean of 4 trials performed by each mouse, per day of learning. ‡: p < 0.05 vs. 12 m P (M), Two-way ANOVA with repeated measures (Days x Treatment) and Sidak’s multiple comparisons test. Latency to target was also analyzed to determine in each group at what day the mice started to remember the localization of the hidden platform: * p < 0.05 vs. Day1, Two-way ANOVA with repeated measures (Days x Treatment) and Dunnet’s test. (E, F, G) Percentage of time spent in the target quadrant (TQ, quadrant in which the platform was hidden) and the opposite quadrant (OQ) by male (M) and female (F) mice after treatment with placebo (P) or ABT-263 (ABT) at 6-month-old (6 m, E) or 12-month-old (12 m, F) during the probe test. This probe test was performed on day 5, 1 h after the last acquisition of the learning phase (1 h memory retention, E, F, G) and 72 h after the end of the learning phase (delayed memory retention, H, I, J). *: p < 0.05 vs. TQ; Two-way ANOVA with repeated measures (Quadrant x Treatment) and Sidak’s multiple comparisons. Discrimination index (%) reflecting 1 h memory retention (G) and 72 h memory retention (J) in all groups of mice. * p < 0.05, Two-way ANOVA (Sex x Treatment) and Sidak’s multiple comparisons test. N = 15 in each group (placebo and ABT-263, males and females). Data are expressed as mean ± SEM of n mice. *: p < 0.05; **: p < 0.01; ***: p < 0.001; ****: p < 0.0001
In 12-mo male mice, when compared to placebo, the curative treatment with ABT-263 accelerated the learning process on D4 (p = 0.08) and significantly on D5 (p < 0.05), the latency time to reach the target was twofold shorter in 12-mo ABT-263 (15 ± 4 s) than in 12-mo placebo mice (33 ± 6 s, p < 0.05) (Fig. 2C). In female mice, in contrast, the learning curves were similar between 12-mo placebo (D5: 20 ± 4 s) and 12-mo ABT-263 (D5: 24 ± 4 s) treated groups (Fig. 2D). However, whereas 12-mo placebo female mice started to learn to locate the platform as soon as D2 (12-mo placebo D2: 25 ± 5 s vs. D1: 38 ± 4 s, p = 0.0109), in 12-mo female mice treated with ABT-263, this learning process was delayed by 3 days (12-mo ABT-263 D5: 24 ± 4 s vs. D1: 39 ± 3 s, p = 0.0297) (Fig. 2D). These data suggest that the curative treatment with ABT-263 significantly improved the learning process in middle-aged ATX male mice but was deleterious in females. Mean swimming speed over the 5 days of the learning process was not affected by sex or treatment (Figure S3), ruling out any locomotor bias.
One hour after the last test of the learning curve (on D5), the platform was removed and mice were subjected to short-term memory retention probe test: the time spent in the target quadrant (TQ; quadrant where the platform was located) and in the opposite quadrant (OQ) was measured (Fig. 2E, F, G). The same probe test was then repeated 72 h later, to assess delayed memory retention (Fig. 2H, I, J). From these data, a discrimination index for short (1 h) and delayed (72 h) probe tests was calculated (the higher the index, the better memory retention; Fig. 2G, J). In young male mice, ABT-263 significantly improved both short and delayed memory retention (Fig. 2E-G, H-J); in young females, ABT-263 only tended to improve delayed memory retention (p = 0.098, Fig. 2J). Of note, short-term discrimination index in 6-mo placebo females tended (p = 0.065) to be higher than in 6-mo placebo males (males: 28 ± 17% vs. females: 59 ± 12%) (Fig. 2G), suggesting that there is little room for improvement in females with ABT-263 in prevention; in other words, at 6-mo, placebo female mice tend to have better short-term memory retention than placebo males.
In middle-aged mice, curative treatment with ABT-263 improved (p < 0.05) short-term memory retention in males but had no effect in females (Fig. 2F-G). As observed in younger mice, short-term discrimination index in 12-mo placebo females was higher than in 12-mo placebo males (males: 23 ± 18% vs. females: 63 ± 8%, p = 0.075) (Fig. 2G), suggesting again that there is little room for improvement in females with ABT-263. ABT-263 tended to improve delayed discrimination index (p = 0.183) in males, but not in females (Fig. 2J); nevertheless, the time spent in the TQ was significantly different than that in the OQ (p < 0.05), only in ABT-263-treated male mice, not in placebo-mice, suggesting significant delayed memory retention with the treatment (Fig. 2I). In contrast, in 12-mo females, whereas in placebo-treated mice TQ time was significantly different than OQ time (p < 0.05), in ABT-treated mice, the time spent in TQ was similar than that in OQ (p > 0.05) (Fig. 2I), suggesting that ABT-263 was deleterious in females. A significant interaction between the variables “sex” and “treatment” in the 2-way ANOVA of the discrimination index illustrates an opposite effect of ABT-263 in male and female mice: for short-term discrimination index F(1, 54) = 6.700, p = 0.0124 and for delayed discrimination F(1, 54) = 4.259, p = 0.0439. Mice used to assess the effects of ABT-293 on cognition were tested before and after the treatment, i.e. mice were pre-trained, which could affect cognitive responses at the end of the treatment. However, according to our data (not shown), pre-training had very limited effects, if any, on cognitive response to ABT-263 in ATX mice. Altogether, these data suggest that both preventive and curative treatments with ABT-263 improved various aspects of learning and memory retention only in male ATX mice.
Impact of ABT-263 on atherosclerotic lesions and cerebrovascular flow-mediated endothelium-dependent dilationATX mice develop spontaneously an aortic atherosclerotic plaque under a regular diet [40, 42]. Cerebral arteries are plaque-free (data not shown). Aortic lesions increased with age, from 6- to 12-mo, similarly in male and female ATX mice (Fig. 3A).
Fig. 3
Impact of ABT-263 treatment on atherosclerotic lesion (A) Aortic lesion (% area) was quantified by oil-red-O in 6 m (n = 19 males and n = 15 females), 9 m (n = 22 males and n = 23 females) and 12 m (n = 32 males and n = 36 females) ATX mice. *: p < 0.05 vs. 6 m, one way-ANOVA and Dunnett’s multiple comparisons test. (B, C) Top panels, typical Oil-Red-O-colored thoracic aortas with atheroma plaques; bottom panels, atherosclerotic plaque expressed as percentage of total aortic area of both males (M) and females (F) mice treated with placebo (P) or ABT-263 (ABT) at 6-month-old (6 m, n = 19–21 males, n = 17–18 females, B) or 12-month-old (12 m, n = 26–28 males, n = 27 females C). *: p < 0.05 vs. Placebo, Two-way ANOVA (Treatment x Sex) and Sidak’s multiple comparisons test
The preventive treatment with ABT-263 had no beneficial effect on aortic lesions in 6-mo males, but in females, ABT-263 paradoxically increased lesion area when compared to placebo (+ 33%, from 7 ± 1 to 9 ± 1%, p < 0.05) (Fig. 3B). In contrast, curative treatment with ABT-263 in middle-aged mice had no effect in female mice but slightly reduced the size of the lesion by 15% (from 39 ± 1 to 33 ± 2%, p < 0.05) in male mice (Fig. 3C).
Cerebral endothelium-dependent dilation to shear stress was globally unaffected by age and was similar in males and females (Fig. 4A). In males, the preventive treatment improved endothelium-dependent dilatory responses to shear stress, with no significant beneficial effect in females (Fig. 4B). Indeed, in 6-mo male mice, ABT-263 increased sensitivity to shear stress (i.e., decreased EC50; from 7.6 ± 0.8 in placebo to 5.7 ± 0.4 dyn/cm2 in ABT-263, p < 0.05) but not efficacy (no modification of Emax) (Fig. 4C). In contrast, the curative treatment had no effect in 12-mo males but highly improved efficacy of shear stress on endothelium-dependent dilatory responses in 12-mo females (i.e., increased Emax from 36 ± 4 to 59 ± 5%, in placebo and ABT-263, respectively; p < 0.05) (Fig. 4D).
Fig. 4
Impact of ABT-263 treatment on cerebral endothelial dilatory function (A) Flow-mediated endothelium dependent dilations were measured ex-vivo in pressurized pial cerebral arteries from 6 m (n = 13 males and n = 12 females), 9 m (n = 16 males and n = 17 females) and 12 m (n = 18 males and n = 22 females) ATX mice. (B, C, D, E) Flow-induced dilation of cerebral arteries of both male (M) and female (F) mice treated with placebo (P) or ABT-263 (ABT) at 6-month-old (6 m, n = 16 males, n = 11–12 females, B) or 12-month-old (12 m, n = 15–18 males, n = 16–17 females, D). EC50 (sensitivity to shear stress: the higher the sensitivity, the lower EC50) of cerebral arteries of male or female mice treated with placebo or ABT-263; Emax (efficacy to shear stress, the higher the efficacy the higher Emax) of cerebral arteries of male or female mice treated with placebo or ABT-263 at 6-month-old (6 m) (C) or at 12-month-old (12 m) (E). *: p < 0.05 vs. Placebo, Two-way ANOVA (Sex x Treatment) and Sidak’s multiple comparisons test. Data are mean ± SEM of n mice
Altogether, these data suggest that ABT-263 effects on atherosclerotic plaque size were disconnected from the partial improvement of cerebrovascular endothelial dilatory function, in both male and female mice.
Opposite sex-dependent effect of ABT-263 on cerebrovascular SASP markersIt is not clear whether ABT-263 crosses or not the BBB [59], and thus whether the senolytic lowers or not senescence in the brain. We quantified the effects of ABT-263 on the cerebrovascular expression of p21 and different SASP factors (Fig. 5 and Table 1): in young 6-mo mice, preventive treatment with ABT-263 significantly reduced IL6 expression both in male and female mice (Fig. 5), but otherwise had little effect on the SASP markers. However, the cerebrovascular expression of p21 (a marker of cell cycle arrest [60, 61]), IL6, PAI1, TNFα and Angptl2 (4 members of the SASP [60,61,62]) were higher in females than in males, in placebo and/or ABT-263 groups (Fig. 5 and Table 1), highlighting a sexual dimorphism. Indeed, a main sex-effect was observed for these markers in young mice: p21 (F(1, 28) = 4.485, p = 0.0432), PAI1 (F(1, 26) = 16.19, p = 0.0004), Angptl2 (F(1, 28) = 14.10, p = 0.0008), TNFα (F(1, 26) = 14.75, p = 0.0007) (Fig. 5 and Table 1).
Fig. 5
Effect of ABT-263 treatment on gene expression of cerebrovascular p21 and SASP markers. (A) p21 (B) IL6, (C) PAI1, and (D) Angptl2 expression in male (M) or female (F) mice treated for 3 months with placebo (P) or ABT-263 (ABT) at 6-month-old (6 m) or 12-month-old (12 m). Data are mean ± SEM of n (n = 7–8) mice. *: p < 0.05 vs. Placebo, #: p < 0.05 vs Males, ANOVA-2way (Sex x Treatment) followed by Sidak’s multiple comparisons. Main sex-effect (#) and main treatment-effect (*) are also indicated when the interaction between the 2 independent factors was not significant. *: p < 0.05; ****: p < 0.0001; #: p < 0.05; ##: p < 0.01
Table 1 Impact of preventive (from 3 to 6-mo) and curative (from 9 to 12-mo) ABT-263 treatment on gene expression in the cerebrovascular fraction (data are 2^-ΔΔCT)At 12-mo, ATX mice exhibit an increased BBB permeability previously demonstrated by Evans blue extravasation and by the presence of microbleeds [1], which could permit diffusion of ABT-263 into the parenchyma [50]. In the cerebrovascular fraction, gene expression of markers of senescence-associated damage tended to decrease in 12-mo males (p21, −32% p = 0.107; IL6, −52% p = 0.116, Angptl2, −25% p = 0.166;) (Fig. 5), without significant impact on the parenchymal fraction (Table 2). In contrast, in 12-mo females, ABT-263 paradoxically increased p21 (p21, + 73% p < 0.05) and SASP factors of inflammation in the cerebrovascular fraction (PAI1, + 53% p < 0.05, IL6, + 125% p = 0.085, and Angptl2 + 34% p = 0.107) (Fig. 5); ABT-263 also increased two recognized markers of senescence in the parenchymal fraction in females (p21 + 302%, p < 0.05 and LMNA, + 40% p = 0.058) (Table 2). Altogether, these data suggest an opposite sex-dependent effect of ABT-263 on cerebrovascular p21 and SASP factors, illustrated by significant interactions between the two independent variables “sex” and “treatment”: vascular p21 (F(1, 27) = 12.22, p = 0.0017), Angptl2 (F(1, 26) = 7.176, p = 0.0126), PAI1 (F(1, 28) = 4.776, p = 0.0374), IL6 (F(1, 28) = 8.297, p = 0.0075) (Fig. 5), and parenchymal p21 (F(1, 28) = 6.55, p = 0.0162) and parenchymal LMNA (F(1, 27) = 7.170, p = 0.0125) (Table 2). The rise in p21 and SASP markers in 12-mo ABT-263-treated female ATX mice could, at least partially, be associated with the absence of improved cognition in female mice. Of note, in contrast to p21, the senescence marker p16 was not detectable in the brain of ATX mice, in any of the groups, neither at the gene (cerebrovascular or parenchymal fractions), nor at the protein level (hippocampal region) (data not shown).
Table 2 Impact of preventive (from 3 to 6-mo) and curative (from 9 to 12-mo) ABT-263 treatment on gene expression in the parenchymal fraction (data are 2^-ΔΔCT)Opposite sex-dependent effect of ABT-263 on endothelial cells densityBoth preventive and curative treatments with ABT-263 increased CD31-positive endothelial cells density in the hippocampus area of male mice; in females, ABT-263 had the opposite effect in young mice and no effect in middle-aged mice (Fig. 6). Cerebrovascular ICAM1 mRNA expression was also significantly decreased (p < 0.05) by ABT-263 in 6-mo females (Table 1). In addition, the expression of claudin-5 coding for a tight junction at the BBB, was increased by ABT-263 in young 6-mo male mice (p = 0.065), but significantly decreased (p < 0.05) in females: main sex-effect for claudin-5 (F(1, 28) = 4.496, p = 0.0430). In middle-aged mice of both sexes, ABT-263 had no impact on claudin-5 or occludin (Table 1). Altogether, these data suggest a beneficial effect of ABT-263 on endothelial cells markers in male mice, but not in females.
Fig. 6
Effect of ABT-263 treatment on hippocampal protein marker of endothelial cells. Immunostaining in the hippocampus of CD31, a marker of endothelial cells in males (M) and females (F) ATX mice treated with placebo (P) or ABT-263 for 3 months (ABT), at 6-month-old (6 m) or 12 month-old (12 m). Typical images and a negative control for each staining are shown; scale bar = 100 µm, for all images. Data are mean ± SEM of n (n = 15–18) images (% area of positive staining / % area of DAPI-labelled nuclei). *: p < 0.05 vs Placebo, #: p < 0.05 vs Males, ANOVA-2way (Sex x Treatment) followed by Sidak’s multiple comparisons. *: p < 0.05; **: p < 0.01; ###: p < 0.001
Opposite sex-dependent effect of ABT-263 on astrocytes and glial cells activationIn agreement with an opposite sex-effect on cognition in ABT-263-treated mice, astrocyte activation (GFAP) tended to be reduced in males at 6-mo (p = 0.133) and 12-mo (p = 0.171) but astrogliosis was significantly increased in young females (Fig. 7). Parenchymal GFAP gene expression did not change significantly, highlighting a discrepancy between protein and mRNA expression of GFAP in 6-mo females (Table 2).
Fig. 7
Effect of ABT-263 treatment on hippocampal protein marker of astrogliosis. Immunostaining in the hippocampus of GFAP, a marker of astrocytes in males (M) and females (F) ATX mice treated with placebo (P) or ABT-263 for 3 months (ABT), at 6-month-old (6 m) or 12 month-old (12 m). Typical images and a negative control for each staining are shown; scale bar = 100 µm, for all images. Data are mean ± SEM of n (n = 15–18) images (% area of positive staining / % area of DAPI-labelled nuclei). *: p < 0.05 vs Placebo, #: p < 0.05 vs Males, ANOVA-2way (Sex x Treatment) followed by Sidak’s multiple comparisons. **: p < 0.01; ##: p < 0.01
In addition, IBA1-positive glial cells decreased significantly only in male ABT-263-treated mice, at both ages (Fig. 8), together with lower brain parenchymal Iba1 gene expression in males than in females mice: in 6-mo mice, main sex-effect for Iba1 (F(1, 26) = 16.04, p = 0.0005) and in 12-mo mice, main sex-effect for Iba1 (F(1, 27) = 20.82, p < 0.0001) (Table 2). This suggests decreased microgliosis in ABT-263 treated males. Although IBA1 protein expression was not affected by ABT-263 in young and middle-aged female mice (Fig. 8), Iba1 gene expression was significantly increased by the treatment at both ages in females (Table 2); with these discrepancies between protein and mRNA expression of IBA1, our data suggest either no benefit or increased microgliosis in ABT-263 treated females.
Fig. 8
Effect of ABT-263 treatment on hippocampal protein marker of gliosis. Immunostaining in the hippocampus of IBA1, a marker of glial cells in males (M) and females (F) ATX mice treated with placebo (P) or ABT-263 for 3 months (ABT), at 6-month-old (6 m) or 12-month-old (12 m). Typical images and a negative control for each staining are shown; scale bar = 100 µm, for all images. Data are mean ± SEM of n (n = 15–18) images (% area of positive staining / % area of DAPI-labelled nuclei). *: p < 0.05 vs Placebo, #: p < 0.05 vs Males, ANOVA-2way (Sex x Treatment) followed by Sidak’s multiple comparisons. **: p < 0.01; ***: p < 0.001; #: p < 0.05
Synaptophysin is a pre-synaptic protein that reflects synaptic density. Better cognition in male mice treated with ABT-263 is not matched by an increased synaptic density, as RT-qPCR data in brain parenchymal fraction (in 12-mo mice, not observed in young mice) show that synaptophysin expression was higher in 12-mo ABT-263 females than males, main sex-effect in 12-mo for synaptophysin (F(1, 27) = 8.638, p = 0.0067) (Table 2). Similarly, brain parenchymal BDNF expression was higher in 12-mo female mice than in males, main sex-effect for BDNF (F(1, 27) = 7.829, p = 0.0094) (Table 2). However, expression of the neuronal marker NeuN was not different between groups (Table 2). Altogether, higher Synaptophysin and BDNF expression may reflect a potentially better “memory reserve” in females. Concomitant higher levels of parenchymal inflammatory IL6 (p = 0.066), TNFα (p = 0.111) and Iba1 (p = 0.061) in 12-mo placebo-female mice than in males (Table 2), may have compromised the effect of ABT-263 on cognition in females.
Comments (0)