
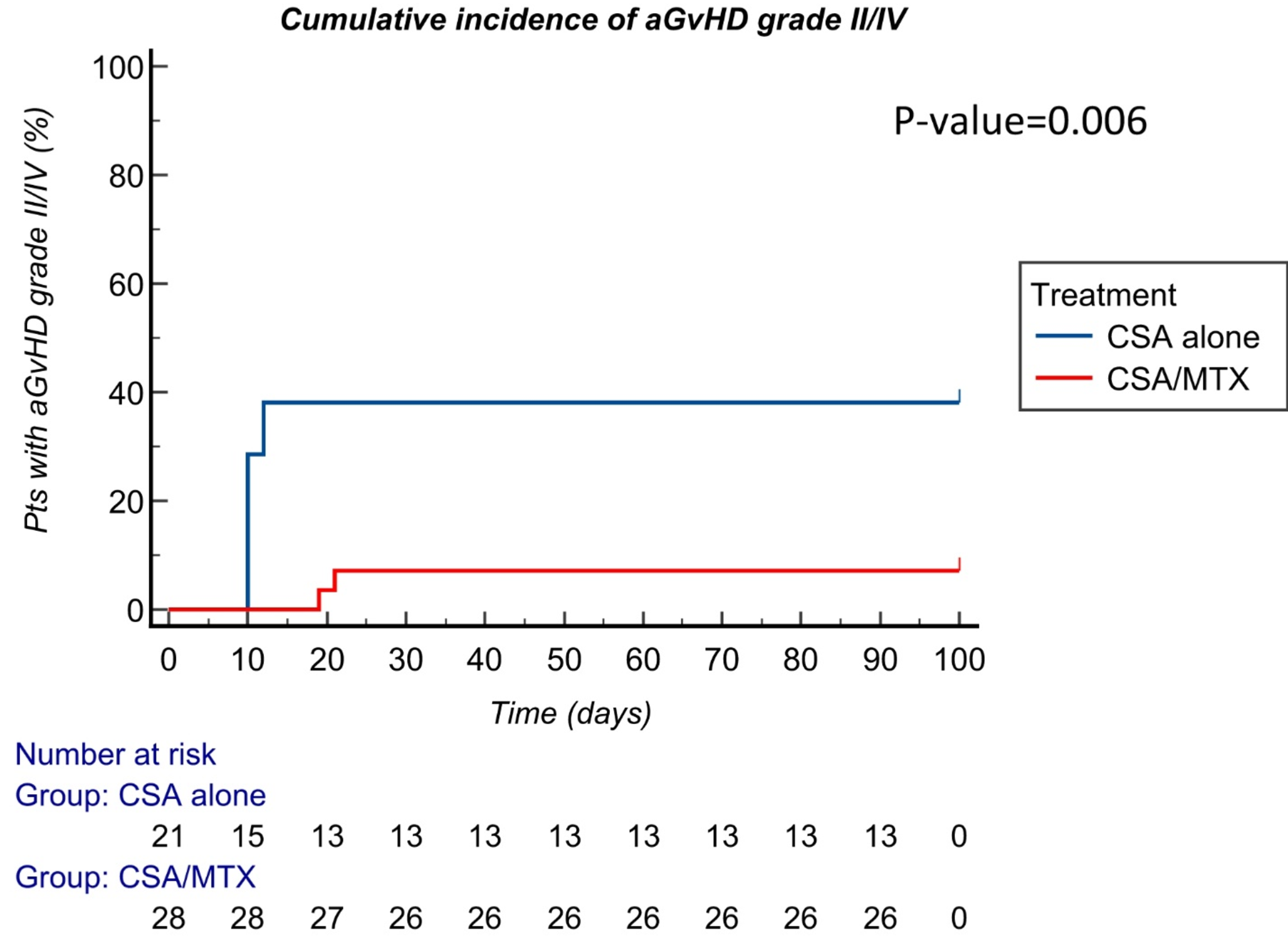
Remember me


In the present study, a comprehensive analysis led to the identification of 501 DEGs. These DEGs were pinpointed through the debatching merge matrix of GSE50161, GSE104291, and GSE122498 (Fig. 1A). Subsequently, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses were conducted on these identified genes. The GO analysis outcomes revealed that they were predominantly enriched in several key biological processes and cellular components, such as ho protein signal transduction, coated membrane, AP-type membrane coat adaptor complex, and chloride channel activity. Meanwhile, the KEGG analysis demonstrated that the target genes were chiefly concentrated in pathways related to nicotine addiction, arginine and proline metabolism, beta-Alanine metabolism, and histidine metabolism (Fig. 2).
Fig. 2
The differentially expressed genes were analyzed by GO and KEGG. The differentially expressed genes were analyzed by GO and KEGG
WGCNA analysisIn the realm of WGCNA, the determination of the soft threshold power represents a linchpin step. This parameter wields substantial influence over the robustness and interpretability of the ensuing gene co-expression network. To ascertain the optimal soft threshold power, network topology analysis was used to ascertain the optimal soft threshold power. After painstaking calculations and iterative model refinements, a soft threshold power of 9 was determined for this particular WGCNA analysis. Significantly, this value corresponds to the minimal power requisite for achieving a scale-free topological fit index of 0.9, as vividly depicted in (Figs. 3A, B). This strategic choice laid the groundwork for a more reliable and biologically relevant network construction.
Fig. 3
WGCNA analysis. A β = 10,0.85; B β = 10,9.33; C 8 important modules; D Interaction between modules
Once the soft threshold power was determined, the research proceeded to the next step. All available genes were used as building blocks to meticulously construct a hierarchical clustering tree. This dendrogram served as a blueprint, carving out eight prominent and functionally distinct modules (Fig. 3C). These modules, akin to interconnected hubs within the gene network, were then subjected to an in-depth exploration of their mutual interactions (Fig. 3D). Unraveling these relationships was crucial in deciphering the underlying genetic regulatory mechanisms.
As documented in the seminal work \*MERGEFORMAT (Tang et al. 2018), state-of-the-art statistical techniques were employed to compute the expression correlation between module feature vectors and genes, yielding the Module Membership (MM) metric. By applying a stringent cutoff of (|MM|> 0.8), 1909 genes with pronounced connectivity within the clinically relevant modules were identified. These genes, acting as the veritable powerhouses of the network, were aptly designated as hub genes. Their identification not only sheds light on the core regulatory circuitry but also paves the way for future investigations into disease pathogenesis and potential therapeutic interventions.
Construction and analysis of protein–protein interaction (PPI) networkTo gain deeper insights into the functional relationships among differentially expressed genes (DEGs), a protein–protein interaction (PPI) network was constructed. A comprehensive PPI network specific to the DEGs under investigation was assembled by utilizing the STRING online database. Subsequently, this network was imported into Cytoscape software for in-depth analysis (Fig. 4A). Two distinct and advanced algorithms were employed in the pursuit of identifying key regulatory elements. These algorithms meticulously sifted through the network’s complex architecture, ultimately pinpointing the hub genes (Fig. 4B, C). Through this rigorous process, a set of 10 core genes, which are likely to play central roles in the biological processes under study, were successfully identified.
Fig. 4
Construction and analysis of protein–protein interaction (PPI) network. A PPI network of DEGs; B, C 10 core genes
Metascape enrichment analysisMetascape, a powerful bioinformatics tool, was harnessed to perform enrichment analysis. The outcomes of this analysis, which encompassed both the pathways and biological processes implicated, are visually presented in Figs. 5 and 6. These figures offer a detailed snapshot of the complex web of molecular events and cellular functions that the DEGs are involved in, providing crucial clues for further research.
Fig. 5
Pathways and processes for Metascape enrichment analysis
Fig. 6
Results of the Metascape enrichment analysis
Core gene expression analysisFor the Gene Set Enrichment Analysis (GSEA), the GSEA software (version 3.0) was sourced directly from its official website (DOI: 10.1073 / pnas. 0506580102, http://software.broadinstitute.org/gsea/index.jsp). In this study, samples were meticulously partitioned into two distinct groups, with one group comprising glioma tumor samples and the other consisting of non-tumor samples. To conduct a comprehensive analysis, The c2.cp.kegg.v7.4.symbols.gmt file was further procured from the Molecular Signatures Database (DOI: 10.1093 / bioinformatics/btr260, http://www.gsea-msigdb.org/gsea/downloads.jsp).
This setup enabled us to evaluate the relevant pathways and molecular mechanisms underlying the studied phenomenon. Specifically, based on the gene expression profiling and the predefined phenotype grouping, we established a series of stringent criteria for statistical significance. A minimum gene set size of 5, a maximum gene set size of 5000, along with 1000 rounds of resampling were implemented. Additionally, only those results with a P value less than 0.05 and a false discovery rate (FDR) lower than 0.25 were considered to carry meaningful biological implications (Fig. 7). Through this rigorous Gene Set Enrichment Analysis (GSEA) process, the aim was to unravel the hidden regulatory patterns and gain a more profound understanding of the gene functions within the context of glioma and its associated biological processes.
Fig. 7 Heat map of gene expression
Heat map of gene expressionTo understand glioma’s molecular differences, the core gene expression was studied. The transcriptional profiles of glioma tumor and non—tumor samples were compared to identify genes with significant expression differences. This helps explain glioma development and find biomarkers or targets.
Expression data was analyzed using advanced techniques and tools. Figure 8 shows clear differences in some core genes. These genes may be key in cell processes like proliferation, apoptosis, and angiogenesis, linked to glioma. Studying their differential expression could clarify glioma biology and lead to better diagnostics and treatments.
Fig. 8
Heat map of gene expression. Differential expression of core genes between glioma tumor and non-tumor samples
Core gene expression analysisThrough meticulous expression analysis, 10 genes crucial for brain glioma (CENPF, PBK, ASPM, KIF2C, KIF20A, CDC20, TOP2A, NUSAP1, TTK, KIF23) were found to be relatively highly expressed in tumor tissues. Statistical significance was achieved with a P-value less than 0.05 (Fig. 9).
Fig. 9
Core gene expression analysis. Ten core genes related to glioma were relatively highly expressed in tumor tissues.A ASPM; B CDC20; C CENPF; D KIF2C; E KIF20A; F KIF23; G NUSAP1; H PBK; I TOP2A; J TTK
CTD analysisIn this study, the CTD website was utilized by inputting the list of hub genes to probe for diseases associated with the core genes, thereby deepening the understanding of gene-disease associations. The 10 genes of focus (CENPF, PBK, ASPM, KIF2C, KIF20A, CDC20, TOP2A, NUSAP1, TTK, KIF23) were found to have potential correlations with multiple medical conditions.
Notably, connections to carcinomas, especially hepatocellular carcinoma, were identified. Dermatological implications were evident, with contact dermatitis being a prime example. There were also links to chemically/drug-induced liver injury and necrosis, suggesting impacts on hepatic cell viability. Hyperplasia, a potential precursor to cancer, was implicated, along with weight loss, potentially due to perturbed metabolic pathways. Inflammation, a ubiquitous factor in diseases, was involved, hinting at immune-modulating capabilities of these genes. Prenatal exposure-delayed effects underlined long-term consequences. Hepatomegaly and neoplasms, including experimental models, were associated, highlighting the genes’ roles in tumorigenesis and organ pathology. Figure 10 visually captures these complex relationships, providing a foundation for further research into mechanisms and potential therapies (Fig. 10).
Fig. 10
Analysis of CTD. A ASPM; B CDC20; C CENPF; D KIF2C; E KIF20A; F KIF23; G NUSAP1; H PBK; I TOP2A; J TTK
Prediction and functional annotation of miRNA associated with hub genesWithin the framework of this research endeavor, The Targetsacan platform was utilized by inputting the list of hub genes to explore relevant microRNAs (miRNAs), aiming to enhance the comprehension of gene expression regulation (Table 1). This investigation revealed specific miRNA associations for key genes. For the CENPF gene, the related miRNAs were identified as hsa-miR-302c-3p.2 and hsa-miR-520f-3p. In the case of the KIF20A gene, its associated miRNA was hsa-miR-153-3p. The TOP2A gene was found to be correlated with hsa-miR-144-3p, while the TTK gene was linked to hsa-miR-455-3p.1. Additionally, for the KIF23 gene, the associated miRNA is hsa-miR-103a-3p. These miRNA-gene relationships offer crucial insights into the intricate regulatory mechanisms governing gene expression, potentially paving the way for future therapeutic interventions.
Table 1 A summary of miRNAs that regulate hub genesImmunofluorescence and western blot analysisImmunofluorescence and western blot analysis elucidated a significant expression pattern in glioma. Notably, CENPF exhibited a high level of expression, while p53 was downregulated (P < 0.05). Concomitantly, both CENPF and CDK-1 were highly expressed, which was accompanied by a diminished expression of p53, p21, and Caspase 9. This coordinated alteration led to the suppression of the apoptotic pathway, consequently fueling glioma progression.
Upon overexpression of CENPF, the key components of the p53-mediated apoptotic pathway were further repressed. In contrast, when CENPF was underexpressed, these pivotal elements of the apoptotic pathway were activated, triggering apoptosis in glioma cells (Figs. 11, 12). Understanding these molecular dynamics provides a profound basis for deciphering glioma pathophysiology and paves the way for potential therapeutic manipulations.
Fig. 11
Immunofluorescence analysis of CENPF
Fig. 12
Western blot analysis. CENPF, CDK-1, p53, p21, Caspase9. P < 0.05
Comments (0)