This study used a subset of data collected for previously published works [8, 9]. Specifically, data from the mice implanted with electrodes in the mPFC, vHPC, and VTA (see Drive implant) were selected to examine the simultaneous interactions between these areas. Unlike the previously published reports demonstrating how novelty exposure enhances subsequent learning [8, 9], the present study focused on the time course of neural dynamics in the mPFC-vHPC-VTA circuit through novelty exposure per se.
Subjects
Three-month-old male C57BL/6J mice (Jackson Labs) were kept on a 12-h light/ 12-h dark cycle with lights on at 7 am. Food and water were available ad libitum. After chronic implant surgery, mice were paired and separately housed in cages divided by a perforated plastic divider to prevent social isolation and protect the implants. Mice were randomly assigned to experimental groups on the day of the experiment. All procedures adhered to the NIH Guidelines and were approved by Columbia University and the New York State Psychiatric Institute Institutional Animal Care and Use Committees (IACUC).
Surgical procedures
Anesthesia was induced at 2% and maintained at 0.8% isoflurane throughout surgery. Mice were placed on a heating pad. Carprofen (0.15 mL) and dexamethasone (0.05 mL) were administered subcutaneously before the surgery. Experiments were performed four weeks after surgery when the mice were fully recovered.
Drive implant
A tungsten wire field electrode (76 μm diameter) was implanted into the dHPC (targeting CA1 pyramidal layer; 1.9 mm posterior to, 1.3 mm lateral to, 1.26 mm below bregma), vHPC (targeting ventral CA1/subiculum; 3.2 mm posterior to, 3.3 mm lateral to, 4.59 mm below bregma), and VTA (3.2 mm posterior to, 0.32 mm lateral to, 4.43 mm below bregma). A bundle of 13 tungsten wire stereotrodes (13 μm diameter) was implanted into the mPFC (targeting prelimbic/ infralimbic cortex, layer II/III;1.8 mm anterior to, 0.3 mm lateral to, 2.1 mm below bregma). The wires were connected to a 36-channel electrode interface board (EIB, Neuralynx), which was fixed to the skull with dental cement. Reference and ground screws were placed in the skull overlying the frontal cortex and cerebellum, respectively and fixed with dental cement. The wires attached to the screws were connected to the EIB.
Cannula implant
Guide cannulae (26 gauge; Plastics One, Roanoke, VA 24018) were implanted bilaterally into the vHPC at a 10-degree angle (3.2 mm posterior to, 3.88 mm lateral to, 3.3 mm below bregma) and secured with dental cement. The dummy cannulae (Plastics One) were inserted into the guide cannulae until the day of drug infusion. A tungsten wire field electrode was attached to a guide cannula with the tip of the electrode positioned 0.7 mm below the cannula. Additional field electrodes were implanted into the dHPC and VTA, and a stereotrode bundle was implanted into the mPFC as described above. On the day of infusion, 33-gauge internal cannulae with a 0.5 mm projection were inserted into the guide cannulae.
Histology
After completing behavioral experiments, electrode placements were confirmed by visual inspection of electrolytic lesions. Mice were anesthetized using a ketamine/xylazine mixture, and lesions were created by passing a 50 μA current to an electrode for 20 s. Mice then underwent transcardial perfusion with PBS followed by 4% paraformaldehyde in PBS. Brains were fixed overnight at 4°C in 4% paraformaldehyde and cryoprotected in 30% phosphate-buffered sucrose at 4°C for three days. Using a cryostat, brains were sectioned into 40 μm slices and mounted with DAPI Fluoromount-G mounting medium (Southern Biotech, Cat. #: 0100–20). Only data from validated recording sites were included in data analyses.
Drug
The selective D1-like antagonist SCH23390 (Tocris, Cat. #: 0925) was prepared as a 100 mM stock solution in saline and delivered at 1 mM final concentration in saline.
Behavior
Previous reports indicate that exposing mice to a brightly lit open arena induces anxiety-related behavior and increases vHPC-mPFC theta synchrony. Notably, these effects were not observed when the same experiments were performed in the dark [29]. Moreover, introducing bright light during novel arena exposure abolishes novelty-induced synaptic depression [30]. Therefore, all experiments were performed in the dark to avoid any non-specific anxiety-related effects on brain circuitry.
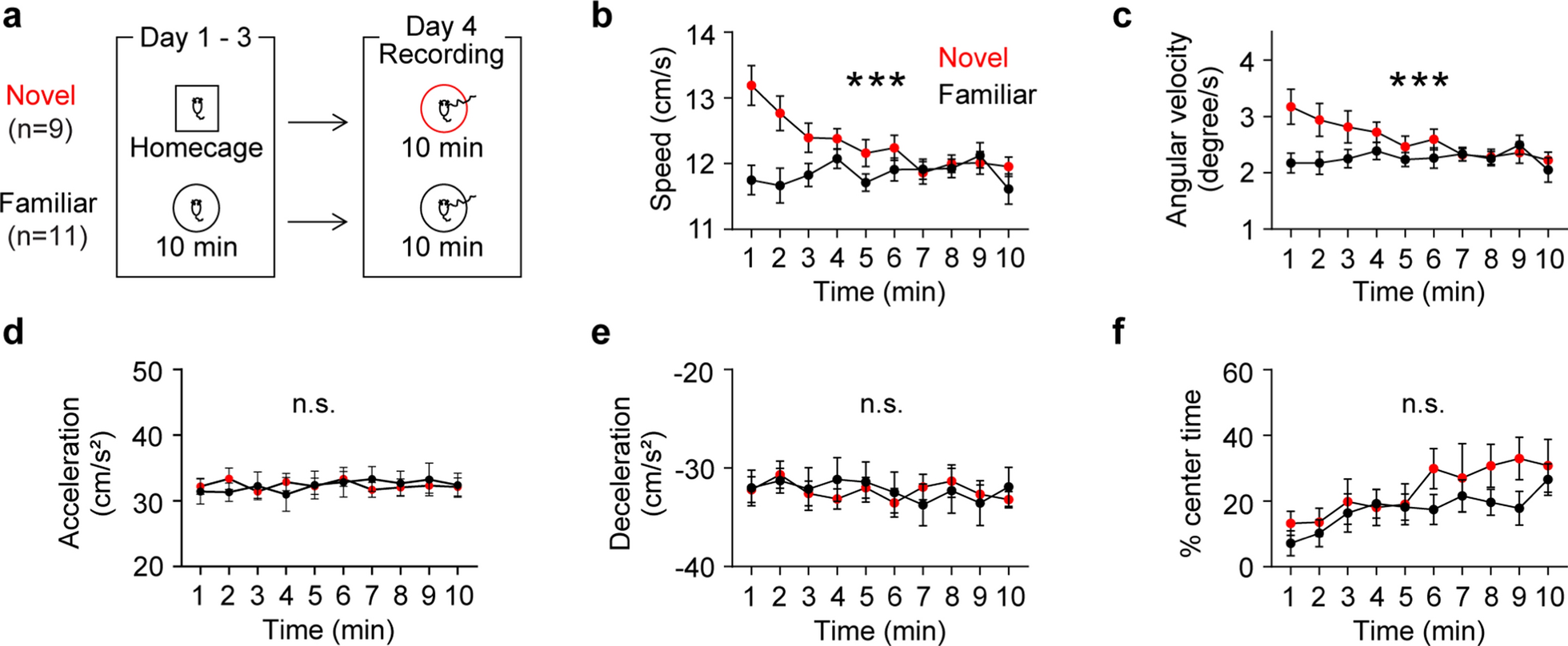
Four weeks post-surgery, food restriction began to maintain 85% of the pre-restriction weight until the end of the experiment. On the same day, a three-minute gentle handling was started and continued for three days to acclimate mice to the experimenter. Then, for the next three days, a subset of mice (the familiar group) was exposed to a circular arena (50 cm in diameter and 25 cm in height) for 10 min to allow them to be familiarized with the arena. The rest of the mice (the novel group, the cage mates of the familiar group) remained in their home cage. Finally, the day after the last day of arena exposure of the familiar group, both groups of mice were exposed to the same arena for 10 min.
D1 receptor antagonist experiment
Mice were habituated to cannulation procedures for five days prior to the day of infusion to minimize potential novelty associated with the procedures. SCH23390 (100 nl, 1 mM) or vehicle (saline, 100 nl) was loaded into a 10 μL Hamilton syringe and administered bilaterally to the vHPC at 50 nl /min using a Harvard Apparatus Pump II Dual Syringe micropump. Following administration, the injection cannulae remained in place for five minutes to let the injected fluid diffuse. After twenty minutes in homecage, mice were introduced to the novel circular arena. Of note, although injecting 3.1 mM of SCH into the dHPC suppresses novelty-enhanced memory consolidation [8], injecting such a high concentration into the vHPC leads to severe sedation or bradykinesia for several hours. The injected 100 nl of 1 mM SCH23390 is equivalent to 0.1 µmol/kg, which is the minimum effective concentration that acts on D1 receptors [31]. SCH23390 is also an agonist for 5-HT2 receptors, but the minimum effective concentration for these receptors is 30 µmol/kg [31]. Therefore, although it cannot be ruled out, it is unlikely that the observed effect of SCH23390 treatment is due to the activation of 5-HT2 receptors.
Neural data analysis
Neurophysiological recordings were conducted while mice were exploring the circular arena. A Digital Lynx system (Neuralynx, Bozeman, MT) was used to amplify, band-pass filter (1–1000 Hz) and digitize the electrode recordings. LFPs were sampled at 2 kHz. In order to mitigate the effect of animal movement [32], recordings were analyzed when mice were moving (6–30 cm/s). This range was determined based on the observed bimodal distribution of speeds during the exposure [8]. All analyses were performed when both groups of mice were moving within the same range of speed and acceleration. Data were analyzed using custom-written MATLAB scripts. Raw LFP data were normalized to the root mean square of the voltage signal for the entire session to address impedance variability across electrodes. Power and coherence were calculated using the wavelet method (MATLAB wavelet toolbox). Phase information was extracted using the Hilbert transform to obtain theta phase angles. Phase angle differences between two LFP signals were calculated as follows:
$$Phase\,synchrony = \left| \,\sum\limits_^ - \phi_ } \right)}} } } \right|$$
n is the number of time points. Pi is a phase angle from brain area x or y at time t.
Statistics
Graphpad Prism 9 was used for statistical analysis. All statistical tests were two-tailed. Differences were considered statistically significant when P < 0.05. All data passed normality tests (Anderson–Darling and Shapiro–Wilk), allowing the use of two-way RM ANOVA tests. Non-parametric tests were also used for within- and between-group analyses of LFP data.




Comments (0)