
Remember me
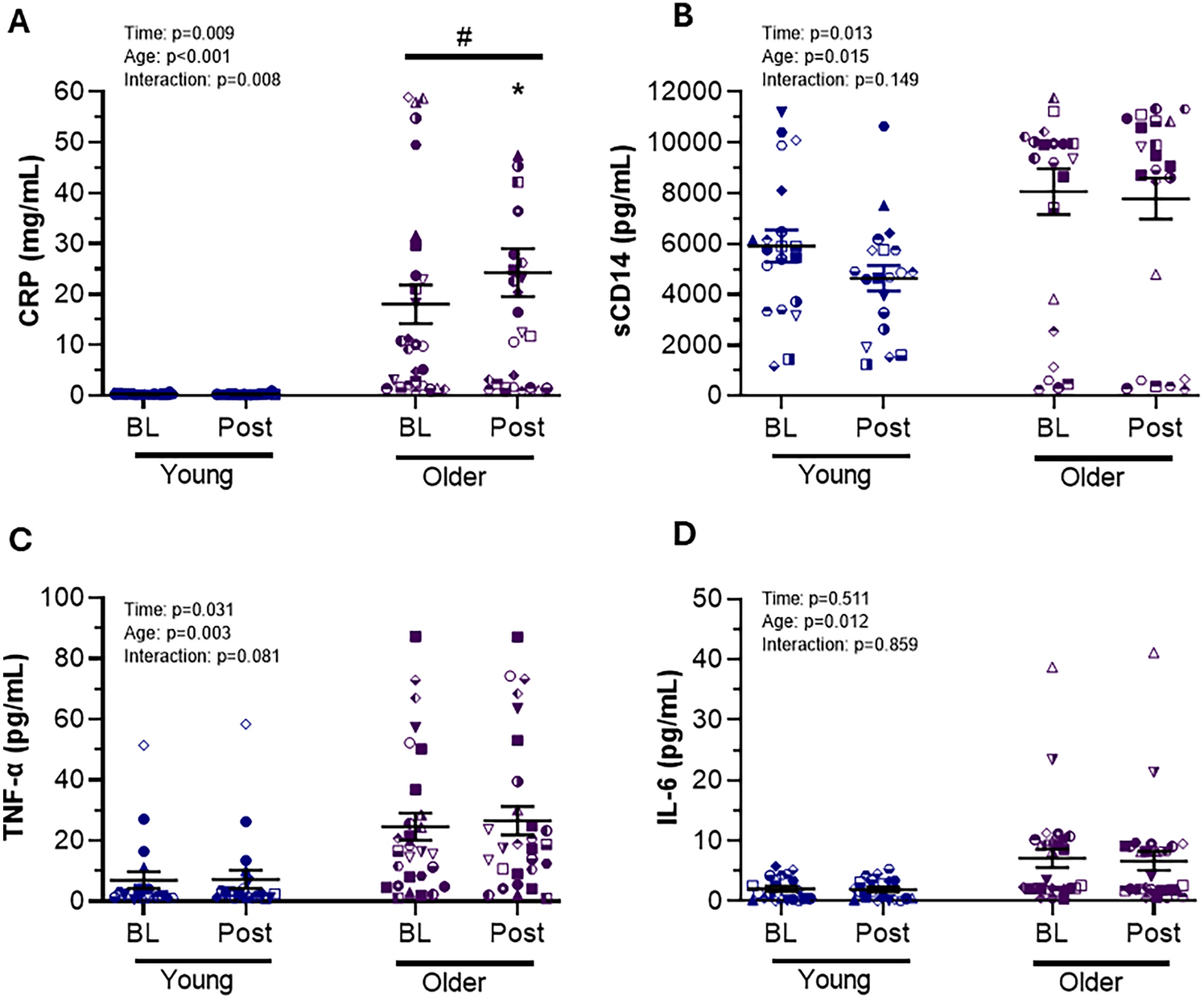
This study investigated the effects of load carriage in normoxia and normobaric hypoxia on ventilatory responses, hemodynamics, tissue oxygenation, and metabolism. Primary findings of this study included the following: (1) load carriage depresses the VT and SV response to exercise matched for absolute VO2 in normoxia; (2) increases in Q and VE during normoxic load carriage matched for walking speed and hypoxic load carriage matched for absolute VO2 or speed were primarily accomplished via increases in HR and fB, respectively; (3) normoxic load carriage increased cerebral O2HHb without influencing muscle oxygenation; (4) muscle HHb was increased with hypoxic load carriage matched for absolute VO2 and walking speed, but muscle O2HHb was only affected when matched for speed; (5) cerebral O2HHb and HHb were reduced and increased, respectively, with load carriage in hypoxia; and (6) load carriage did not independently influence substrate utilization when matched for relative and absolute intensities; however, hypoxia increased CHO dependence. The following discussion will focus on matched absolute VO2 and walking speed stages for cardiorespiratory responses owing to the predictable nature of findings for exercise of a lower oxygen demand (i.e., matched relative intensity). Discussion of oxygen kinetics and metabolic data will include all intensities.
Ventilatory responsesLoad carriage in normoxia induced a shallower breathing pattern when matched for absolute VO2. Specifically, VT and fB were reduced and increased, respectively, relative to U while VE was consistent between normoxic conditions. Comparable alterations in VT and fB have been reported by others investigating the effects of thoracic load carriage (Phillips et al. 2016b; Armstrong et al. 2019), chest wall restriction with inelastic straps (Miller et al. 2002; Tomczak et al. 2011), or fiberglass chest casting (Coast and Cline 2004) and have been attributed to reductions in operating lung volumes (Shei et al. 2017). Few studies have assessed ventilatory mechanics with unloaded exercise compared to load carriage matched for oxygen demand (Phillips et al. 2016b, c; Shei et al. 2018). A study by Phillips et al. (2016b) similarly reported decreases and increases in fB and VT, respectively, with prolonged (45 min; ~ 3.0 L min−1) load carriage (25 kg). However, differences in VT were not observed until late in exercise (i.e., 35 min), which contrasts with the present study (i.e., 10 min stages; 20 total min of exercise at stage 2 end). Additional distinctions occurring late in exercise were increases in VE and VO2 with load carriage, which the authors suggested were likely the result of ventilatory compensation for increases in deadspace ventilation.
An explanation for these discrepancies with Phillips et al. (2016b) is not immediately clear but may be related to methodological differences relating to exercise intensity and ventilatory demand. In a study by Dominelli et al. (2012), the effects of carrying various loads at two fixed workloads on operating lung volumes and the power (i.e., energy requirement) of breathing were investigated. With increasing loads up to 35 kg, end expiratory lung volumes were reduced. Interestingly, the power of breathing was maintained with the heaviest load relative to no load when ventilation was matched at ~ 45 L min−1 but increased substantially at higher levels of VE (~ 70 L min−1). This finding suggests that the alterations in operating lung volumes or breathing mechanics induced by the heavy load may have been compensatory adaptations permitting the maintenance of the power of breathing and VE at the lower ventilatory demand. However, this altered breathing pattern may have been insufficient to meet ventilatory needs at higher workloads, thus requiring adoption of breathing patterns more consistent with unloaded exercise (e.g., similar or higher VT) despite the apparently necessary increase in the power of breathing to achieve it. This theory is supported by ventilatory data from the present study in which a shallower breathing pattern was adopted at approximately the same “low” VE (~ 45 L min−1; matched absolute VO2 stage) while a similar VT was observed between U and LN when matched for walking speed, an intensity necessitating a higher VO2 and VE (~ 70 L min−1) in the loaded condition. Importantly, potential increases in the power of breathing at high VE rates may contribute to fatigue of the respiratory muscles during prolonged exercise and deterioration of breathing mechanics over time (e.g., increases and decreases in fB and VT, respectively), thereby requiring increases in VE to offset reductions in alveolar ventilation. This hypothesis is supported by the data from Phillips et al. (2016b), which employed exercise resulting in a VE of ~ 75 L min−1, which caused respiratory muscle fatigue in the loaded condition and increases in VE over time likely to compensate for reductions in VT late in exercise. More research is clearly warranted to investigate these questions and determine whether a potential threshold exists for VE-dependent alterations in breathing mechanics that influence the power of breathing.
The predictable increases in VE with hypoxic load carriage relative to unloaded normoxic exercise were primarily achieved via increases in fB. Specifically, when matched for absolute VO2, VT and fB were maintained and increased, respectively, relative to U. At the same walking speed, fB was further increased relative to U to meet the increased ventilatory demands. To our knowledge, only one study has investigated the ventilatory responses to load carriage in acute hypoxia. In this study, Hinde et al. (2018) assessed breathing mechanics and respiratory muscle fatigue with load carriage (5.5 km; 18.2 kg) in hypoxia (~ 11.8% FIO2; ~ 4300 m) or cold (− 10 °C). Similar to the present results for normoxic load carriage, reductions and increases in VT and fB, respectively, were observed with load carriage relative to unloaded walking. Interestingly, this effect spanned environmental conditions. However, this study did not control for intensity, allowing subjects to self-select walking speeds, which, while beneficial for ecological validity, prevents easy interpretation of ventilatory responses owing to differences in VO2 across conditions. Nevertheless, careful examination of available data from this study in combination with results from the matched oxygen demand and walking speed stages of the present study seem to lend further support for the notion that ventilatory demands may mediate breathing mechanics during load carriage and that this theory also applies to hypoxic conditions. Specifically, Hinde et al. (2018) reported relatively low VE values (≤ 30 L min−1) across all conditions and time points coupled with apparent trends for shallower breathing patterns (which may have achieved statistical significance given greater statistical power), in the hypoxic conditions with load vs. unloaded walking at sea level (e.g., 0.82–0.83 vs. 0.87–0.98 and 35–41 vs. 28–34 for VT and fB, respectively). In the present study in which VE at matched oxygen demand and walking speed was ~ 55–95 L min−1 in the hypoxic condition, VT was either similar or increased relative to U and LN, which again suggests that an increased power of breathing may have been required to achieve this higher VE. Importantly, this potential elevation in ventilatory work would likely hasten the development of respiratory muscle fatigue and the attendant metaboreflex (St Croix et al. 2000; Derchak et al. 2002), particularly in hypoxia where diaphragm fatiguability is increased (Babcock et al. 1995; Reinhard et al. 2023). Given this, more research is needed to assess how hypoxia, ventilatory demand, and breathing mechanics may interact to influence respiratory fatigue and blood flow responses.
Hemodynamic responsesThoracic load carriage in normoxia altered hemodynamic variables in ways that mirrored ventilatory responses. Specifically, SV and HR were reduced and increased, respectively, while Q was maintained with LN compared to U at the same absolute VO2. Miller et al. (2002) observed similar responses for SV and HR when chest wall restriction was induced via inelastic strapping. However, this study also reported a 12% reduction in Q, which was not observed in the present study. It is likely that differences in the degree of chest wall restriction contributed to these conflicting findings for Q. Indeed, inelastic strapping has been reported to reduce total lung capacity and forced vital capacity by 33% and 40%, respectively (Miller et al. 2002; Tomczak et al. 2011), whereas load carriage of comparable mass to the present study reduced forced vital capacity by only 4–12% (Dominelli et al. 2012; Phillips et al. 2016b; Armstrong et al. 2019) and did not affect total lung capacity (Phillips et al. 2016b). This suggests attenuated chest wall restriction with thoracic load carriage, which may permit maintenance of Q. Studies investigating hemodynamic responses to thoracic load carriage report equivocal findings, which are likely due to methodological differences (i.e., type and mass of load, subject characteristics) or confounding variables (i.e., environmental conditions) (Sagiv et al. 2006; Nelson et al. 2009).
Our finding of reduced EDV with LN vs. U at matched oxygen demand suggests attenuated venous return as a likely mechanism for the SV reduction with load. Venous return is influenced by the respiratory muscle pump, which creates negative pressure swings via alternating decreases (i.e., inspiration) and increases (i.e., expiration) in intrathoracic pressures (Miller et al. 2005). Attenuated negative pressure swings have been reported with chest wall restriction via inelastic straps (Miller et al. 2002) and could conceivably impact venous return with thoracic load carriage. Alternatively, reductions in SV may have resulted from increases in sympathetic activation and consequent increases in SVR stemming from a potential respiratory muscle metaboreflex. Indeed, others have reported a metaboreflex-induced increase in SVR with increased respiratory muscle work (Sheel et al. 2001, 2002). Moreover, a number of load carriage studies have observed elevations in arterial blood pressure suggesting elevated sympathetic outflow (Hong et al. 2000; Sagiv et al. 2006; Ribeiro et al. 2014). While blood pressure was not measured in the present study, our finding of similar SVR and muscle oxygenation (discussed below) between U and LN at matched absolute VO2 suggests that a respiratory muscle metaboreflex was less likely to be a contributing mechanism. Indeed, Sheel et al. (2002) observed that, with graded increases in inspiratory muscle work, elevations in limb vascular resistance only occurred with contractions sufficient to elicit diaphragm fatigue, which may not have occurred in the present study given the nature/degree of respiratory muscle work (i.e., resulting from thoracic load carriage) and relatively short windows of exercise surrounded by periods of rest.
Interestingly, SV and EDV were maintained and SVR reduced with LH compared to U at matched VO2 and walking speeds. An increase in SV with exercise in hypoxia relative to normoxia at these intensities would generally be expected (e.g., with unloaded exercise) as part of the normal cardiovascular compensation for reduced oxygen availability (Naeije 2010). Moreover, reductions in SVR are logical given the vasodilatory response in active muscle to the increased metabolite production (i.e., from higher relative intensities in hypoxia) and oxygen demand (i.e., at matched speed) at these intensities (VanTeeffelen and Segal 2003; Segal and Bearden 2012). Nevertheless, the matching of SV between U and LH is somewhat unexpected given the previously described reduction in SV with load carriage in normoxia. Moreover, prior studies have observed increases and decreases in sympathetic outflow and leg blood flow, respectively, during hypoxic exercise with inspiratory resistance (Katayama et al. 2013) or hypoxemic exercise with resistive breathing (i.e., among COPD patients) (Simon et al. 2001), which would likely increase afterload. While purely speculative, it is possible that any potential respiratory muscle pump-mediated mechanism influencing venous return was abolished by the maintenance of a similar breathing pattern (i.e., matched VT) and thus respiratory pump effectiveness with LH compared to U. Alternatively, the relatively short exercise duration may have prevented development of respiratory muscle fatigue sufficient to induce a metaboreflex that could affect SVR. Regardless of the precise mechanism, our findings suggest that load carriage does not compromise Q in either environmental condition when matched for oxygen demand, but it is yet to be determined how hemodynamic responses may be altered by more prolonged load carriage in hypoxia where respiratory muscle fatigue is more likely to occur.
Oxygen kineticsTo our knowledge, this is the first study to investigate the effects of thoracic load carriage in different environmental conditions on oxygen kinetics. In muscle during normoxic exercise, oxygenation and regional blood flow was similar between unloaded and loaded exercise. Additionally, HHb was reduced with LN compared to the other conditions at matched relative intensity likely owing to less oxygen extraction given the lower oxygen demand. Based on the aforementioned potential for blood flow redistribution via a respiratory muscle metaboreflex (St Croix et al. 2000; Derchak et al. 2002) or the increased use of accessory muscles attendant to load carriage (Holewijn 1990; Devroey et al. 2007), it was predicted that locomotor muscle oxygenation would be reduced by load carriage. These results indicate that load carriage does not impair oxygen delivery to locomotor muscle during exercise. However, more research is needed that assesses blood flow and oxygen kinetics with unloaded and loaded exercise matched for higher intensities in which central factors may be more limiting to oxygenation and exercise performance.
Muscle oxygenation was also mostly maintained with load carriage in hypoxia except at the highest exercise intensity (i.e., matched speed). Additionally, oxygen extraction appeared to be elevated at the higher intensities as evidenced by increases in HHb. Similar levels of muscle oxygenation combined with increased oxygen extraction at matched workloads have been reported by some (Subudhi et al. 2008; Rosales et al. 2022), but not others (Subudhi et al. 2007; Angeli et al. 2019), with hypoxic vs. normoxic exercise. An explanation for this equivocal data may be found in the methodological differences between studies, which include exercise modality, duration, hypoxia type (i.e., normobaric or hypobaric), and muscles assessed. Another interpretation of the present study’s findings, when accounting for the within-condition reduction in oxygenation with LH between matched relative and absolute intensities, and the between-condition reduction with LH compared to the other conditions at matched speed, is that muscle oxygenation seemed to progressively decline with intensity. Such an intensity-dependent reduction in muscle oxygenation has been reported previously (Subudhi et al. 2007, 2008), and may be mediated by regional blood flow. Indeed, tHHb concentrations were increased during LH at matched relative intensity compared to the other conditions, which likely contributed to the aforementioned maintenance of tissue oxygenation. Thereafter, tHHb was similar between conditions, which may have resulted in insufficient blood flow to sustain oxygenation levels (i.e., given likely lower oxygen diffusion and arterial oxygen content) requiring increased oxygen extraction to compensate. Thus, our data indicates that regional blood flow may be limiting for muscle oxygenation at moderate to high intensities in hypoxia, but not normoxia. Of interest, prior studies have reported progressive increases in muscle tHHb with exercise intensity during cycling in either normoxia or hypoxia (Subudhi et al. 2007). In the present study, tHHb did not change during exercise in the LH condition despite progressive increases in intensity. As such, it is tempting to speculate that this may be a load carriage-specific response. However, further study comparing unloaded vs. loaded exercise in hypoxia is needed to confirm this.
Cerebral oxygenation responses indicate a load carriage-specific effect in normoxia. Specifically, cerebral oxygenation was increased at every exercise intensity with load carriage while HHb was similar or lower compared to U. An explanation for this interesting finding is not immediately clear. It seems reasonable to assume that increased oxygenation is the result of increases in blood flow, which might be expected with LN vs. U given: (1) the higher relative exercise intensities and consequent increased sympathetic activation/blood pressure (i.e., at matched oxygen demand) (Hong et al. 2000; Sagiv et al. 2006; Ribeiro et al. 2014) and (2) the increased Q (i.e., at matched speed). Our data mostly supports this notion, as we noted a strong trend (p = 0.051) for an increase in tHHb at matched oxygen demand and a statistical increase in tHHb at matched speed. As previously discussed, it is possible that load carriage results in blood flow redistribution stemming from chest wall restriction or increased respiratory muscle work. It is conceivable that these effects may augment cerebral blood flow in some way. However, more research is clearly needed to elucidate the mechanism for this seeming over-compensation in cerebral oxygenation.
Responses to load carriage in hypoxia for cerebral oxygenation were, in some ways, the inverse of those observed during LN. Specifically, O2HHb was decreased relative to U or LN at rest and every exercise intensity, and HHb was correspondingly increased. In the case of LH, this seemed to be more clearly mediated by increases in cerebral blood flow, as tHHb was increased vs. U or LN at every exercise intensity. This exaggerated tHHb response with LH may be related to impaired cerebral autoregulation in hypoxia combined with the aforementioned load carriage factors affecting blood flow distribution (Derchak et al. 2002; Miller et al. 2005; Ainslie et al. 2007). Regardless of the mechanism, it appears that load carriage increases cerebral blood flow and that changes in hemoglobin status seem to be mediated by environmental condition. Based on prior studies, this is a novel finding. Others have similarly reported reductions and increases in cerebral O2HHb and HHb, respectively, with exercise in hypoxia (Ainslie et al. 2007; Bourdillon et al. 2014; Rosales et al. 2022). However, none of these studies have observed increased tHHb in hypoxia relative to normoxia. It is possible that the observed increase in tHHb was region-specific and not an indication of increases in overall cerebral blood flow. Indeed, others have reported discrepancies between middle cerebral artery blood flow velocity and tHHb in the frontal cortex region (Ainslie et al. 2007; Bourdillon et al. 2014). An additional possibility is that, given the increased biomechanical difficulty of bearing a heavy load (Attwells et al. 2006), frontal cortex motor activity and metabolic demand were increased relative to unloaded exercise thereby enhancing the distribution of cerebral blood flow to that region (Delp et al. 2001), without substantially altering overall cerebral blood flow. Whatever the case may be, our finding has potentially important health implications as cerebral pressure/perfusion has been implicated in the pathophysiology of acute mountain sickness and high-altitude cerebral edema (Hackett 1999; Taylor 2011). Thus, the present study indicates that load carriage exercise may exacerbate the risks for these conditions; although, more study is needed to confirm this.
Metabolism and perceptual responsesThis is the first study to evaluate metabolic responses to load carriage in different environmental conditions and account for the effects of load on relative and absolute exercise intensities. With LN, relative and absolute CHO oxidation were similar to U at the same relative intensity and absolute VO2. This aligns with prior studies (Phillips et al. 2016b, c), which reported similar RER values between loaded and unloaded conditions matched for oxygen demand in males and females. When matched for walking speed, LN increased relative/absolute CHO oxidation and lactate vs. U, which also aligns with prior studies (Blacker et al. 2009; Arcidiacono et al. 2023) and is predictable given the higher relative and absolute exercise intensity in the loaded condition (Romijn et al. 1993). Collectively, these data suggest that load carriage does not independently mediate substrate utilization when matched for relative intensity and oxygen demand with unloaded exercise (i.e., at least given the relatively low matched intensities [40–45%VO2max] employed in this study) but increases CHO dependence when walking at the same speed.
In hypoxia, absolute and relative CHO oxidation were reduced and maintained, respectively, with LH compared to the normoxic conditions at matched relative intensity. This finding aligns with a recent meta-analysis that evaluated the effects of hypoxia on substrate utilization (Griffiths et al. 2019b). Moreover, it was predictable given the lower oxygen demand and overall substrate oxidation with LH at this intensity. When matched for oxygen demand or walking speed, relative/absolute CHO oxidation and lactate were increased compared to the normoxic conditions. This was also predictable given the higher relative intensities imposed by hypoxia during these stages (~ 60–80 vs. 40–45%VO2max) and aligns with prior studies evaluating unloaded exercise (Lundby and Van Hall 2002; Young et al. 2018). Additionally, blood glucose was elevated during LH at all intensities. Prior studies have observed increased rates of glucose appearance and utilization by exercising skeletal muscle in hypoxia (Brooks et al. 1991; Roberts et al. 1996). This glucose dependence in hypoxia has been attributed to the fact that CHO is a more energy efficient fuel than lipids (i.e., yields more energy per liter of oxygen; Mazzeo 2008) and that epinephrine concentrations are increased with hypoxic exposure likely elevating liver glycogenolysis (Mazzeo et al. 1991).
Interestingly, the bioenergetics of walking did not appear to be affected by load carriage despite the above-described cardiorespiratory responses. Specifically, the energy cost of walking (i.e., net above resting) at the same absolute VO2 (~ 1.7 L min−1) equated to ~ 4.0 J kg−1 m−1 across conditions (i.e., when accounting for differences in mass [U = 81.7 kg; LN/LH = 111.7 kg], and speed [U = 1.2 m s−1; LN/LH = 1.0 m s−1 assuming an energy equivalent of 20.9 kJ L−1 O2−1). This value is essentially equal to that reported previously with unloaded walking at a similar speed (~ 1.2 m s−1) and gradient (10%) (Minetti et al. 2002). Collectively, this suggests that walking biomechanics and efficiency are maintained with load carriage in normoxia and hypoxia. However, it is unknown whether this still applies at different speeds, grades, and degrees of hypoxia. A study by Phillips et al. (2016a) observed that VO2 relative to total mass was similar between unloaded and loaded (25 kg) conditions during constant speed (1.5 m s−1) graded exercise testing up to a gradient of 4%, but small increases in oxygen cost occurred in the loaded condition at higher gradients (6–8%). Given this, it is possible that load carriage imposes a narrower range of speeds or relative intensities (Boffey et al. 2019) for optimal biomechanical or metabolic efficiency given different terrain and conditions. Further study is clearly warranted to define these parameters to accurately prescribe paces for populations with occupational load carriage requirements in various environments.
Finally, VO2max and RPE were decreased and increased, respectively, in both loaded conditions relative to U. Smaller magnitude reductions in VO2peak (2.5–3.5%) vs. the present study (7.3%) have been reported previously during graded exercise tests in normoxia with load carriage of similar weight (25 kg). While the reduction in aerobic capacity may be related to any of a number of mechanisms influenced by load carriage/chest wall restriction, the reason for the discrepancy in magnitude between studies is unknown. It is possible that the graded exercise test protocol employed by these prior studies resulted in greater local muscle fatigue (i.e., which we observed during pilot testing of a similar protocol), which prevented achievement of a true max in the loaded condition; but this is purely speculation (Phillips et al. 2016b, c). Reductions in VO2max in hypoxia were expected. However, as in normoxia, the magnitude reduction was slightly larger (~ 33%) than has been reported at similar altitudes/levels of hypoxia previously reported with unloaded exercise (26–27%; Buskirk et al. 1967; Cymerman et al. 1989). While it is possible that this is due to load carriage-specific effects as were observed in normoxia, more research is needed to confirm this. Increases in RPE with load carriage have been reported elsewhere with exercise matched for absolute VO2 (Phillips et al. 2016b, c) and walking speed (Blacker et al. 2009; Armstrong et al. 2019), which are logical given the resultant higher relative and absolute intensitie
Comments (0)