
Remember me
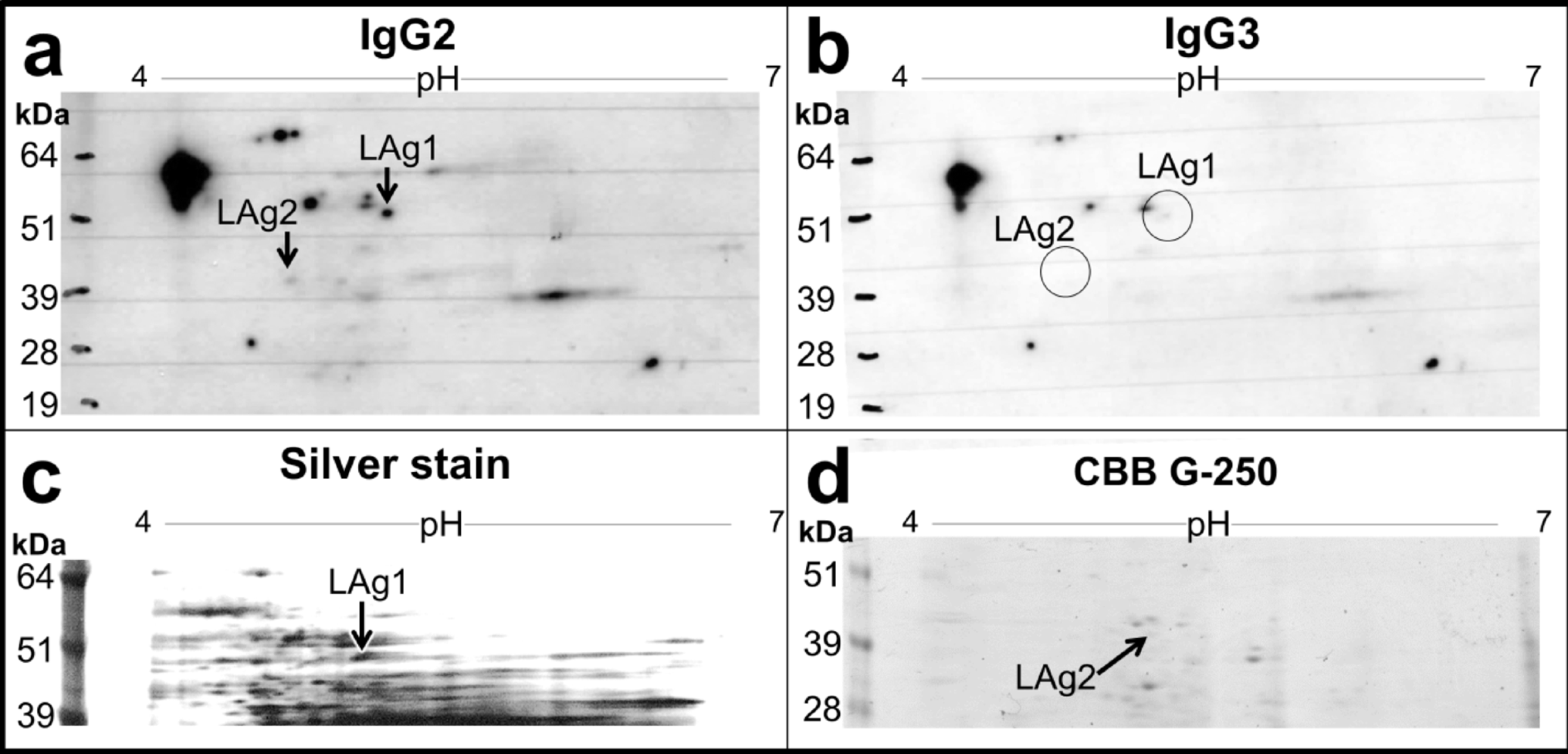
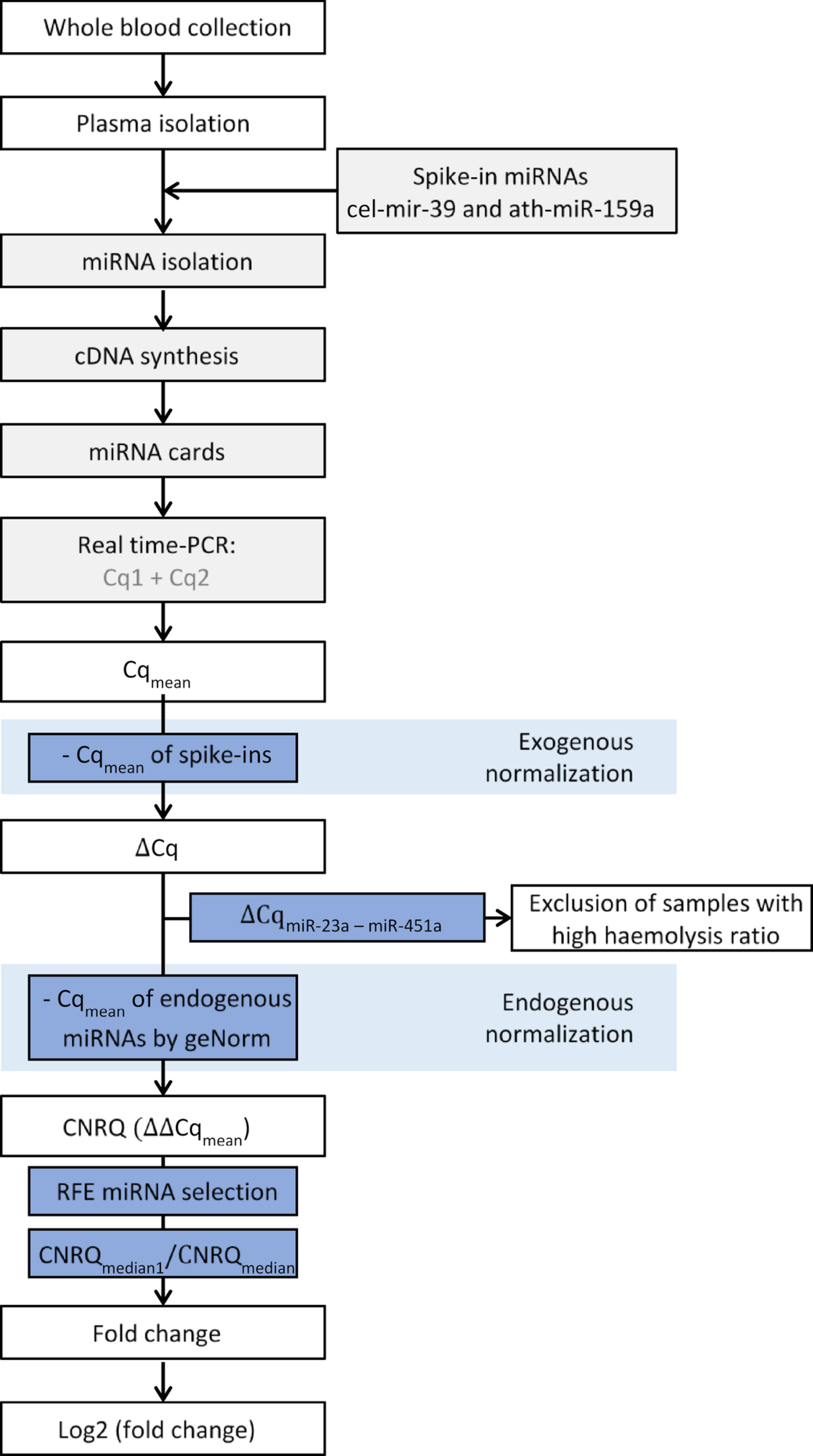
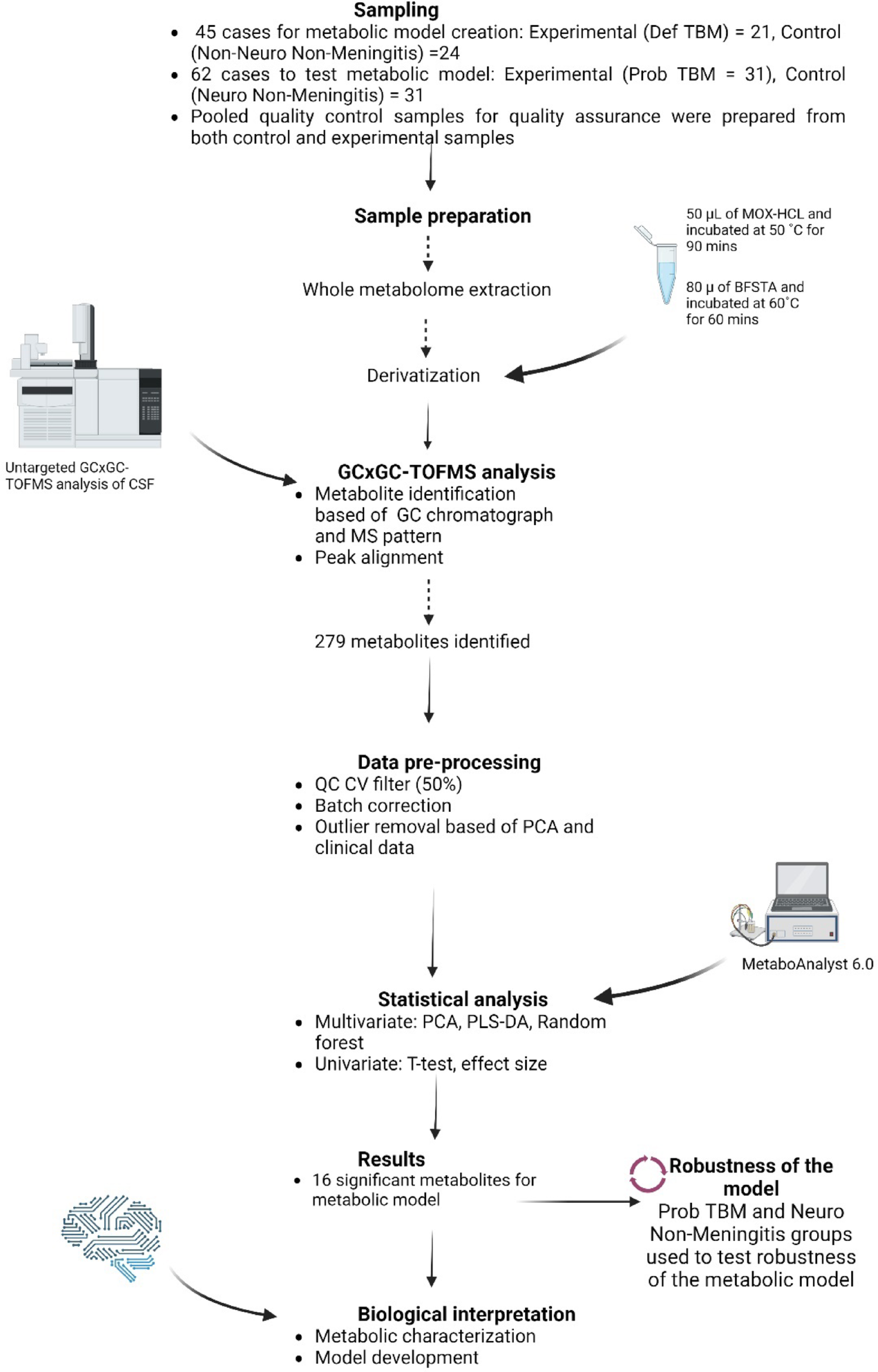
For this study, CSF from 24 control and 21 Def TBM cases were analyzed via GCxGC-TOFMS and underwent statistical analyses. For a metabolite to be considered significant (for characterization purposes), it had to conform to the following cutoff criteria: p-value < 0.05, d-value > 0.5, and VIP > 1.0. A total of 16 significant metabolites were identified in this study (Table 2). In addition to these 16 significant metabolites, 32 metabolites had a d-value > 0.5 and a VIP > 1.0 but did not have a p-value < 0.05 and are also described in Table 2. Metabolites discussed are found in glucose metabolism (galactose; erythrono-1,4-lactone; ribonolactone; tartronic acid), amino acid metabolism (protein serine and 2-ketoisocaproic acid), fatty acid metabolism (undecylenic acid and palmitic acid), imines (3-oxaoct-4-en-2-imine), volatile organic compounds (2-ethylhexanol and 3,5,5-trimethylhexan-1-ol), and various alkenes (1-butene, 1-butoxy-1 butene; 1-pentene;2-(2-methoxyethoxy) acetic acid), amines (5-ethyl-1,2,4-oxadiazol-3-amine) and glycosides (methyl galactose). The metabolic roles of these metabolites in TBM are described below and illustrated as a metabolic model in Fig. 5.
Fig. 5
Schematic of a metabolic model describing M.tb infection in the brain. Pathways affected are color-coded, where light blue indicates the Krebs cycle, pink indicates fatty acid metabolism and volatile organic compounds (VOCs), brown shows the branched-chain amino acids, purple represents the pentose phosphate pathway, and yellow indicates glycolysis. The red arrows represent downregulation of metabolites by TBM, and the green arrows show upregulation. List of abbreviations: AA, amino acids; ADP, adenosine diphosphate; ATP, adenosine triphosphate; STPK, serine/threonine protein kinases; ROS, reactive oxygen species
Metabolic burstThe presence of elevated levels of lactate (lactic acid), sucrose, and 2-(2-methoxyethoxy) acetic acid, accompanied by reduced levels of glucose and galactose, is all indicative of a metabolic burst—an increase in energy production in the M.tb-infected brain, previously described in TBM [7, 12,13,14]. The significantly reduced levels of glucose, accompanied by increased levels of lactic acid, also support the hypothesis of the astrocyte-microglia lactate shuttle (AMLS) [15]. Briefly, glucose levels in the brain decrease during M.tb infection because of increased glycolysis in astrocytes. The glycolytic end-product pyruvate is converted to lactic acid, which is then shuttled from astrocytes to the microglia—the resident immune cell of the brain. Hence, brain energy metabolism becomes focused on fighting the M.tb infection and protecting neurons.
Reduced concentrations of tartronic acid, ribonolactone, and erythron-1,4-lactone in the CSF also support the concept of metabolic burst in TBM patients. Ribonolactone (also known as D-ribono-1,4-lactone) is the oxidized form of ribose and erythrono-1,4-lactone (also known as D-erythronolactone), which is derived from erythose. Ribose and erythrose are both involved in various biological processes including the pentose phosphate pathway and glucose metabolism, where they are used in nucleotide and amino acid synthesis [16]. During glucose metabolism, ribose-5-phosphate and erythrose-4-phosphate can be converted to glucose-6-phosphate, which is then catabolized via the glycolytic pathway, thereby contributing to the metabolic burst. Tartronic acid is a lactate dehydrogenase inhibitor known to block glycolysis, and it also inhibits cytosolic malate dehydrogenase [17, 18]. Thus, reduced tartronic acid in the M.tb-infected brain can result in an increased metabolic burst.
Lastly, significantly elevated levels of 2-ketoisocaproic acid were detected in the CSF of the TBM patient group. 2-Ketoisocaproic acid is formed during the catabolism of leucine (a branched-chain amino acid), serves as an intermediate for the synthesis of glutamate, and can also be utilized in the Krebs cycle for energy production. Hence, significantly increased 2-ketoisocaproic acid further supports the metabolic burst in TBM.
Calcium signalingThis study found two volatile organic compounds (VOCs)—2-ethylhexanol and 3,5,5-trimethylhexan-1-ol, significantly reduced to below detectable concentrations in the TBM group, compared to that in the Non-Neuro Non-Meningitis group. Studies showed that 3,5,5-trimethylhexan-1-ol has been found to induce calcium signaling and cAMP in cell lines, where intracellular calcium is dependent on the cAMP-dependent signaling pathway [19]. M.tb has been shown to inhibit calcium signaling within the body, and it’s this inhibition that results in susceptibility to M.tb infection, as well as M.tb survival and growth [20]. Intracellular calcium also plays a role in autophagy by activating the serine/threonine kinase enzyme, which in turn leads to the phosphorylation of beclin-1 and VPS34, inducing autophagy [21]. Autophagy serves to contain M.tb infection by preventing the fusion of the phagosomes with the lysosomes [22]. Considering this, the inhibition of calcium signaling by M.tb in turn results in the inhibition of these VOCs. Additionally, protein serine was found to have significantly reduced concentrations (non-detectable) in the TBM group compared to that of the Non-Neuro Non-Meningitis cases (0.0007 ± 0.0008 µmol/L). Protein serine concentrations in TBM are indicative of the serine/threonine protein kinases (STPKs) found in M.tb, which act as part of the mechanism for signal transduction, allowing for growth/survival in the host [23]. Thus, the significantly decreased concentrations of 2-ethylhexanol, 3,5,5-trimethylhexan-1-ol, and protein serine in the TBM group support the hypothesis of M.tb-inhibited calcium signaling; however, this is completely speculative at this point and requires future validation studies.
Moreover, it should be noted that VOCs, such as alkanes, are a natural component of the myelin sheath in neurons [24]. Lipid peroxidation, common in TBM, induces alterations in the physical properties of myelin membranes, releasing VOCs [25]. In a study on multiple sclerosis (MS) [26], a disease known for degradation of the myelin sheaths, exhaled VOCs were identified as potential markers of MS (i.e., degradation of the myelin sheath releases VOCs). In fact, some Mycobacterium species have been shown to directly consume alkanes as a source of carbon [27,28,29]. Hence, we postulate that the complete absence of some VOCs in the CSF of TBM cases could be due to the demyelination of neurons, contributing to neuronal damage that is typically associated with TBM.
Fatty acid and alkene metabolismSignificantly reduced concentrations to below the levels of detection for undecylenic acid (UDA) and palmitic acid were identified as key metabolites in this study in the TBM group compared to the Non-Neuro Non-Meningitis group. UDA has been shown to have a neuroprotective effect against neural apoptosis induced by glutamate, hydrogen peroxide, and amyloid beta protein [30]. UDA forms part of the phospholipid layers found in cell membranes of the brain and acts by reducing µ-calpain activity (a type of calcium-activated cysteine protease enzymes where µ-calpain are activated by an influx of calcium ions) [30,31,32]. Caplins are known to play a role in apoptosis by cleaving and activating pro-apoptotic proteins, such as p35, which has an adverse impact on neuron structure and function, potentially leading to neurological decline [30]. Hence, it is likely that the lack of UDA in the TBM group could be an indication of increased neurological deficits associated with TBM due to the decreased caplin activity brought on by the decreased UDA levels. Palmitic acid contributes 20–30% of the total fatty acids found in membrane phospholipids and plays a crucial role in membrane function by assisting with cell division, intracellular membrane movement, signal transduction, and influencing the physical properties of the membrane. Changes in fatty acids are also shown to alter the fluidity of membranes, which has previously been associated with several immune disorders and neurological diseases [33]. Previous research also shows that M.tb utilizes host-derived lipids, including palmitic acid, for replication using beta-oxidation [34].
Unique to our metabolomics investigation, we are reporting altered alkene markers in the CSF of TBM patients. Grant et al. [35] showed that alkenes, such as 1-pentene and 1-butene, are formed during fatty acid metabolism via various enzymatic reactions. The enzymes involved in this process use hemoproteins, including cytochrome P450, which are typically involved in oxygenations. Cytochromes (CYP1B1 and CYP2B) are expressed in the blood–brain barrier (BBB), where they have been shown to have neuroprotective effects on the brain by working in conjunction with the efflux transports in the brain [35, 36]. Other research has also shown that in neurons, cytochromes are involved in neurotransmitter synthesis [37].
The significantly reduced concentrations of fatty acids (used as a substrate for M.tb growth and replication) and alkenes in the TBM group, and the subsequent alteration of the cellular membranes in the brain, are likely a further contributor to the neurological symptoms seen in TBM patients.
Imine, amines, and methyl galactosideIn this study, 3-oxaoct-4-en-2-imine and 5-ethyl-1.2.4-oxadiazol-3-amine also occurred in significantly reduced concentrations in the TBM group compared to the Non-Neuro Non-Meningitis group. Limited research exists describing these compounds; however, quinone imines have been previously associated with cytotoxicity, redox reactions, and ROS formation in cancer in various in vitro studies [38, 39]. Biernacki et al. [40] described 1,2,4-oxadiazoles as having anticancer, antimicrobial, and anti-inflammatory functions. Other research has highlighted that the derivatives of 1,2,4-oxadiazoles exhibit antitubercular action, specifically against M.tb [41, 42]. Preclinical research in fecal metabolomics has also shown that imines are upregulated in mice after oral ingestion of the probiotic Lacticaseibacillus rhamnosus IDCC 3201 and that this probiotic also has neuroprotective properties [43]. Hence, imines in the CSF may also serve in a neuroprotective capacity and/or function in energy production through redox reactions. The reduced levels of these amines and imines in the CSF of TBM patients are likely due to their inhibition/depletion due to the M.tb infection and subsequently contribute to the neurological symptoms seen in these patients.
Methyl galactoside concentrations were significantly reduced in the CSF of the TBM group. Current research indicates that glycosides have a neuroprotective effect, where they can modulate neuroinflammation by targeting the BBB to reduce neuroinflammation and by decreasing the expression of inflammatory markers in glial cells [44, 45].Thus, the depletion of methyl galactoside in the TBM group may be brought on by M.tb, thereby contributing to the neurological complications associated with TBM. Additionally, this could be a potential therapeutic target for the management of TBM.
Testing the robustness of the metabolic markers and their possible diagnostic useFigure 5 is a schematic that explains the 16 significant metabolites and some of the other identified metabolic of this study, in terms of metabolic pathways. Figure 5 is thus a metabolic model describing M.tb infection in the brain from this study. To test the robustness of this metabolic model, we analyzed the CSF derived from participants diagnosed as Prob TBM and the Neuro control group. Based on the results (PCA—Fig. 4), the two control groups overlap with each other, the two TBM groups overlap with each other, and there is a distinct separation between the control and TBM groups. This means that our metabolic model, which describes both probable and definite TBM similarly, is unique from clinical neurological symptoms (such as seizures—febrile, epilepsy, and viral encephalopathy). We recommend further investigating the significant metabolites detected in this study in the CSF of TBM patients for diagnostic applications.
Further inspection of the 9 metabolites that are only present in the TBM groups (Table S2) shows that three metabolites (2,3,4-trihydroxybutanoic acid; 2-desoxy-pentos-3-ulose and glyceric acid) were present in all samples across all stages of TBM. Of note, 2-ketoisocaproic acid and L-proline were significantly lower (if not absent) in the stage 1 TBM samples, arabinofuranose levels were lower in stage 2a samples only, 5-aminoimidazole was low in samples with stage 2b, and for stage 3 samples, 2-(2-methoxyethoxy)acetic acid levels were completely depleted. This suggests further application of these metabolite markers for not only aiding in diagnosing TBM but also for determining the stages of TBM. However, since the number of cases per TBM stage was too small in this study to draw conclusive results, we recommend doing a larger-scale study with more cases (> 20) per TBM stage to investigate the role of metabolites in differentiating TBM stage.
Comments (0)