
Remember me
First, we set out to explore the potential role of Gram-positive anaerobic cocci (GPAC) to induce monocyte activation in blood, using two of the most common bacteriaemia- and sepsis associated GPAC species, P. harei and P. micra, as examples [13]. In addition, E. coli was used as a positive control, given its established role in blood born infections. Specifically, we stimulated whole blood with heat-inactivated bacteria and analysed the monocytes for activation by flow cytometry, analysing activation markers (PDL1, HLA-DR, CD11b and CD14), phosphorylation of members of common signalling pathways (Akt, p38 and NFκB) and intracellular cytokine production (IL-1β, IL-6, IL-8 and TNF) (Fig. 1A). Representative histograms for the data underlying Fig. 1 can be found in Supplemental Fig. 2. At the surface marker level, stimulation of whole blood (n = 5) with heat-killed P. harei resulted in upregulation of PDL1, HLA-DR and CD11b (p < 0.0110) and downregulation of CD14 (p = 0.0004) (Fig. 1B). P. micra induced expression of HLA-DR (p = 0.0029) and CD11b (p = 0.0177) but not PDL1 (p = 0.0824) and downregulation of CD14 (p = 0.0025). Interestingly, P. harei induced a higher expression of PDL1, HLA-DR and CD11b compared to P. micra (p < 0.0309), and a more prominent downregulation of CD14 (p = 0.0110). Finally, as expected, E. coli stimulation induced increased expression of PDL1, HLA-DR and CD11b (p < 0.0191), with a trend of higher expression than both P. harei and P. micra. Additionally, the downregulation of CD14 by E. coli was more apparent than the downregulation induced by P. harei (p = 0.0736) and P. micra (p = 0.0187). At the phosphorylation level, both P. harei (p < 0.0135) and E. coli (p < 0.0196) induced significantly more phosphorylation of all analysed markers compared to the unstimulated control (n = 4) (Fig. 1C). In contrast, P. micra did not induce markable phosphorylation. Accordingly, both P. harei and E. coli showed higher levels compared to P. micra (p < 0.0505), whilst there were only minor differences between E. coli and P. harei. Finally, at the cytokine level (n = 5), E. coli was clearly the most potent inducer of all measured cytokines (p < 0.0001), followed by P. harei which induced significant levels of IL-1β (p = 0.0063), IL-6 (p = 0.0186) and TNF (p = 0.0125), whilst P. micra only induced significant levels of TNF (p = 0.0051) (Fig. 1D). E. coli was accordingly more potent than both P. harei and P. micra (p < 0.0095), whilst P. harei induced more production of IL-1β, IL-6 and TNF (p < 0.0327) compared to P. micra. Taken together, P. harei (and to a much lesser extent P. micra) is a potent inducer of monocyte activation in whole blood.
To confirm that heat-inactivation of the bacteria did not significantly influence our results, we compared the response of live P. harei and P. micra, to investigate if the pattern of activation is similar to heat-killed bacteria using surface markers and cytokine production as readouts (n = 3, Supplemental Fig. 3). Previous studies by de Chateau et al. [18] and ourselves, [24] reported that heat-inactivation of bacteria facilitate handling, whilst having no effect on the bacterial surface proteins. E. coli was excluded from this experimental setup since its doubling time exceeds that of the GPACs by far. The difference at the surface level was less pronounced, but P. harei induced PDL1 expression (p = 0.0064) and P. micra HLA-DR (p = 0.0061, Supplemental Fig. 3A). There was a minor but statistically significant (p = 0.0417) difference in CD14 expression between P. harei and P. micra. However, differences were more pronounced at the cytokine level with marked production of IL-1β (p = 0.0029), IL-6 (p = 0.0042), IL-8 (p = 0.0040) and TNF (p = 0.0042) of P. harei compared to P. micra (Supplemental Fig. 3B). Hence, P. harei induced an overall stronger response compared to P. micra, in line with was observed for heat-killed bacteria.
Fig. 3
Soluble factors of P. harei are potent inducers of immune cell activation in blood. a Schematic outline of the experimental setup. Whole blood was stimulated with conditioned medium from E. coli, P. harei, and P. micra for 4 h before analysis for (B) surface marker expression, (C) phosphorylation or (D) intracellular production of cytokines by monocytes. Growth medium (GM) was used as a negative control. b Displays surface expression of the three markers of activation (PDL1, HLA-DR and CD11b) on monocytes, as analyzed by flow cytometry. c Summary data on phosphorylated NFkB, p38 and Akt in monocytes following activation with CM for 10 min. d Shows data on the intracellular production of cytokines in monocytes. Gates were set according to the growth medium control (without bacteria). Each dot represents a unique donor (n = 3–4), and data is displayed as mean with SD. Data was analyzed with repeated measures one-way ANOVA with Tukey’s multiple comparisons test. The data was generated from two independent experiments. P < 0.05 was considered statistically significant. Panel A was created using Biorender.com (2025, https://BioRender.com/v52sd2a). Ctrl — Control, GM- growth medium, CM — conditioned medium, MFI – Median fluorescence intensity
Monocyte activation induced by P. harei is more pronounced compared to other Gram-positive bacteriaNext, to put our results in the context of Gram-positive bacteria, we compared the response of P. harei and P. micra to other Gram-positive bacteria (i.e. S. aureus and S. pyogenes). At the surface marker level (n = 5), P. harei induced a higher degree of expression of all markers (PDL1 (p < 0.0299), HLA-DR (p < 0.0403) and CD11b (p < 0.0384)) compared to both S. pyogenes and S. aureus (Supplemental Fig. 4A). Additionally, P. harei induced downregulation of CD14 compared to S. pyogenes (p = 0.0007) and S. aureus (p = 0.0028). There were only minor differences for P. micra, as it induced a higher HLA-DR expression (p = 0.0051) compared to S. aureus, but S. pyogenes induced a higher PDL1 expression (p = 0.0301). At the cytokine level, there was no statistically significant differences. This could possibly be due to the small sample size (n = 3), as there was a trend of higher cytokine production of P. harei compared to the other bacteria (Supplemental Fig. 4B). Hence, P. harei, but not P. micra, is a potent activator of monocytes in blood compared to the other Gram-positive bacteria studied here. Therefore, we focused on the comparison to the most potent activator, E. coli.
Fig. 4
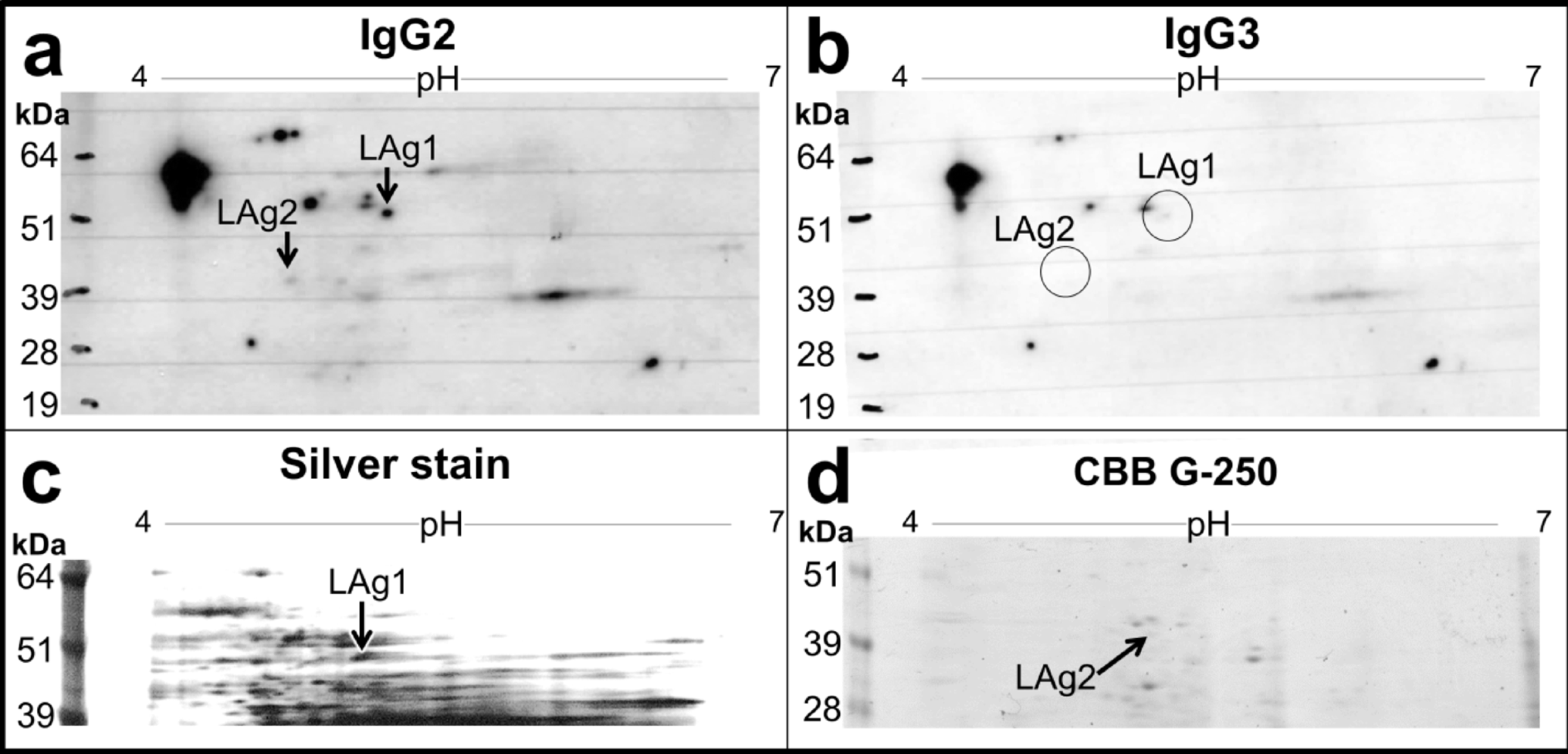
Soluble factors of P. harei induce monocyte activation primarily via TLR2. a Overview of the experimental setup were the mechanism of monocytes activation by conditioned medium of P. harei, was investigates, and potential underlying candidates were explored via gel electrophoresis and mass spectrometry analysis of prominent bands. b Whole blood was pre-treated with anti-TLR2 (10 µg/ml) or anti-CD14 (125 µg/ml) for 1 h followed by addition of supernatants of P. harei. Shows expression of surface markers in monocytes following activation with conditioned medium (CM). Growth medium without bacteria served as negative control and data is presented as fold change (FC) to the negative control. Each dot represents a unique donor (n = 6), and data is displayed as mean with SD. Data was analyzed with repeated measures one-way ANOVA with Tukey’s multiple comparisons test. The data was generated from three independent experiments. Next, P. harei CM was analysed by SDS PAGE and MS. c Gel electrophoresis of P. harei CM. Bands marked with 1–3 were excised and analysed after in-gel digestion by MS and the main identified proteins are highlighted. d STRING analysis of protein networks of the most abundant proteins (coloured balls) found in P. harei CM experimentally determined (magenta lines), database annotated (cyan lines), co-expression (black lines) and gene co-occurrence (dark blue lines) < 10 interactor, confidence 0.500. P < 0.05 was considered statistically significant. Panel A was created using Biorender.com (2025, https://BioRender.com/8lmylbo). MFI — Median fluorescence intensity, CM — Conditioned medium, MS — Mass spectrometry
P. harei activation of monocytes is mainly dependent CD14 signalingNext, we aimed to investigate the mechanism of monocyte activation by P. harei. Several surface proteins of Gram-positive bacteria are known to elicit immune responses, especially lipoteichoic acid (LTA; [25]). Thus, we investigated the expression of LTA in P. micra and P. harei, to check differences in expression. While the previous experiments showed differences between P. micra and P. harei in their ability to trigger immune responses, expression of LTA appeared equal between the species (Fig. 2A). Hence, we instead checked for differences in binding of the bacteria to two different receptors. TLR2 is associated with Gram-positive factor recognition [26, 27] and full TLR2 activation depends on the presence of GPI-anchored CD14 as a co-receptor [28,29,30,31]. We therefore tested the ability of P. harei and P. micra to bind to either receptor. A strong fluorescent signal was detected for the interaction with P. harei with CD14, while no interaction was observed with TLR2 (Fig. 2B, upper panel). Only minor signal was detected for the interaction of P. micra with CD14, as well as for TLR2 (in contrast to P. harei) (Fig. 2B, lower panel). Thus, the pronounced activation of monocytes by P. harei compared to P. micra could be due to more prominent CD14 binding.
To study this, we pre-incubated monocytes (n = 6) with inhibitory antibodies against either TLR2 or CD14, before stimulation with heat-killed P. harei. While only slight changes were detected in the expression patterns with anti-TLR2 pre-treatment, anti-CD14 pre-treatment prevented the increase in PDL1 (p = 0.0067) and HLA-DR (p = 0.0110) by P. harei (Fig. 2C). The trend was similar for CD11b although it was not markedly induced in this experiment. In samples treated with anti-CD14, there was a marked reduction of CD14 expression. This is possibly due to downregulation or recycling of the CD14 receptor as a result of the pre-treatment. Still, we hypothesise that the activation of monocytes in whole blood by P. harei is mainly dependent on CD14 signalling.
Soluble mediators of P. harei are comparable to those of E. coli at inducing monocyte activation in bloodFactors secreted by the bacteria may be a crucial source of virulence and monocyte activation. To investigate a potential effect of soluble mediators, we incubated whole blood with conditioned medium (CM) of the bacteria (Fig. 3A). At the surface level, there was a trend of higher expression of PDL1, CD11b and HLA-DR in monocytes although it did not reach statistical significance, likely due to the data spread and the small sample size (n = 4) (Fig. 3B). Moreover, there was a non-significant trend of lower CD14 expression in E. coli treated samples but not in GPAC treated samples. More strikingly, at the phosphorylation level, both CM from E. coli and P. harei induced phosphorylation of NFκB (p < 0.0272), Akt (p < 0.0055) and p38 (p < 0.0084 (Fig. 3C). Compared to E. coli, there was a trend towards weaker signal in P. harei CM which was significant for Akt (p = 0.0433). No phosphorylation was induced by P. micra CM. Finally, there was a major induction of intracellular cytokine production of IL-1β (p < 0.001), IL-6 (p < 0.0012), IL-8 (p < 0.0043) and TNF (p < 0.0028) in monocytes by E. coli and P. harei, but not P. micra (Fig. 3D). Interestingly, there were no statistical differences in cytokine production between P. harei and E. coli. Taken together, soluble mediators by P. harei potently induce monocyte activation, comparable in several of the markers to that of E. coli.
Monocyte activation by P. harei conditioned medium is dependent on TLR2 signallingSince we observed a positive effect of pre-treating blood with anti-CD14 prior to stimulation with heat-inactivated P. harei, we hypothesised that inhibition of CD14 could also diminish the host responses mediated by the bacterial supernatant (Fig. 4A). However, anti-TLR2 pre-treatment blocked the upregulation PDL1 (p = 0.0005) and CD11b (p = 0.0144) (Fig. 4B). An effect of anti-CD14 could still be observed for PDL1 (p = 0.001) and CD11b (p = 0.0101), although anti-TLR2 inhibition was more potent for PDL1 (p = 0.009). Again, as with heat-killed bacteria, samples pre-treated with anti-CD14 had markedly reduced CD14 expression. Thus, in contrast to the CD14-dependent activation by heat-killed P. harei, activation by P. harei CM is dependent on TLR2 signalling, suggesting different mechanisms of activation depending on interaction with surface bound- or soluble factors.
Identification of possible soluble factors of P. harei involved in monocyte activationNext, we aimed to characterize potential secreted candidates responsible for the observed activation. Gel electrophoresis analysis of the CM identified several bands that were exclusively present in the CM derived from P. harei (Supplemental Fig. 5), while no protein bands were detected in the CM of either P. micra, E. coli or their respective media alone.
Fig. 5
P. harei is a potent activator of neutrophils in blood through similar mechanisms as monocytes. a To generalize our findings, we also investigated the ability of P. harei, P. micra and E. coli to induce neutrophil activation (exemplified by surface marker expression of three activation markers). Heat-inactivated as well as conditioned medium was used to induce activation, and the effects of anti-TLR2 and anti-CD14 were studied using P. harei (C and E). b Whole blood was incubated with heat-inactivated bacteria for 4 h before the analysis of activation markers by flow cytometry (n = 5). The data is presented as median fluorescence intensity (MFI) and depicted as mean with SD. c The blood was pre-treated with anti-CD14 (125 µg/ml) or anti-TLR2 (10 µg/ml) for 1 h before the addition of the bacteria. data is presented as fold change (FC) to the negative control. Each dot represents a unique donor (n = 6), and data is displayed as mean with SD. (d–e) Conditioned medium was used instead of heat-inactivated bacteria to induce activation in n = 4–6 donors. Data was analyzed with repeated measures one-way ANOVA with Tukey’s multiple comparisons test. The data in (B) and (D) were generated from two independent experiments and for (C) and (E) it was generated from three independent experiments. P < 0.05 was considered statistically significant. Panel A was created using Biorender.com (2025, https://BioRender.com/i8bjppr).
To assess whether the detected bands in P. harei CM correlate with known PAMPs, we analysed the CM by SDS PAGE and proteomics mass spectrometry. The three most prominent bands (at > 130 kDa, 110 kDa and 55 kDa; Fig. 4C), were further processed by in-gel digestion mass spectrometric analysis (Supplemental Table 1). These bands were identified based on respective abundances as pyruvate synthase (band 1 at > 130 kDa; calculated molecular weight (MW) 129.6 kDa, Uniprot ID: E4KZ56), SLH-domain-containing protein (band 2 at 110 kDa; calculated MW 99.9 kDa, Uniprot ID: E4KY84) and as putative glycolate oxidase (Uniprot ID: E4L019) or aldehyde dehydrogenase B (Uniprot ID: E4KZR7) (band 3 at 55 kDa; calculated MWs 51.8 and 53.8 kDa, respectively) (Fig. 4C). Whereas the pyruvate synthase, glycolate oxidase and aldehyde dehydrogenase are involved in various biochemical processes, SLH domain-containing proteins have been shown to non-covalently anchor several bacterial proteins to the cell surface [32]; often, proteins that associated with the virulence and survival of various pathogens ([33,34,35]). STRING network analysis of protein interactions (Fig. 4D) revealed that the pyruvate synthase, the putative glycolate oxidase and the aldehyde dehydrogenase interact with proteins involved in bacterial respiration, ion-translocation, fatty acid synthesis and carbohydrate degradation; suggestive of virulence attribute [36, 37]. Taken together, these data highlight several candidates released by P. harei that could be responsible for the observed activation of monocytes in blood.
Immune cell activation by P. harei is not unique to monocytesTo generalize our findings, we next analysed neutrophils in blood (n = 4–6) for activation by using three surface markers (CD66b, CD16 and CD11b) by flow cytometry (Fig. 5A). As with the monocytes, heat-killed P. harei induced activation of neutrophils, evidenced by increased expression of CD66b and CD11b (p < 0.0050), and a downregulation of CD16 (p = 0.0045, Fig. 5B). P. micra also induced activation of neutrophils (p < 0.0129). The magnitude of the response of P. harei was greater than that of P. micra (p < 0.0455) and comparable to E. coli (Fig. 5B). Blood pre-treated with anti-CD14 prevented the upregulation of CD66b (p = 0.0030) and CD11b (p = 0.0072) but did not prevent the CD16 downregulation induced by P. harei (Fig. 5C). Notably, there was no statistical effect of anti-TLR2 pre-treatment.
Using conditioned medium (CM), P. harei induced upregulation of CD66b (p = 0.0306) and CD11b (p = 0.0444), and downregulation of CD16, although it did not reach statistical significance (Fig. 5D). Interestingly, in contrast to the heat-killed bacteria, there was no activation induced by P. micra. Moreover, the activation induced by E. coli was comparable to that of P. harei. However, in contrast to the whole bacteria, we found that the activation induced by P. harei CM was distinctly inhibited by pre-treatment with anti-TLR2 (p < 0.0017, Fig. 5E) in contrast to anti-CD14. Indeed, there was no effect of anti-CD14. Taken together, these results suggest that the activation induced by P. harei is not unique to monocytes, highlighting its potential in inducing inflammation and immune cell activation in blood.
Comments (0)