
Remember me




The primary source of circulating ApoE is the liver.11 To investigate the effect of circulating ApoE on fracture healing we generated mice in which the ApoE gene was selectively nulled in hepatocytes to test whether a loss of hepatic ApoE can improve aged fracture healing. Albumin-Cre mice were crossed with ApoEfl/fl mice to generate hepatocyte-specific knockouts (Alb-Cre+/−;ApoEfl/fl, referred to as ΔApoE) or littermate controls (ApoEfl/fl, referred to as Ctrl) and aged to 24 months (Fig. 1a). ELISA was used on plasma samples to test circulating ApoE concentrations. ΔApoE mice exhibited greater than 90% decrease in plasma ApoE concentration relative to Ctrl (Fig. 1b). Importantly, micro-CT analysis of intact femurs of 24-month-old ΔApoE and Ctrl mice was conducted to determine whether hepatic ApoE affects skeletal development (Fig. S1a, b). No significant differences were identified in cortical thickness, trabecular space, trabecular thickness, nor trabecular number in ΔApoE mice relative to Ctrl (Fig. S1C–F), indicating hepatic ApoE does not affect skeletal development.
Fig. 1
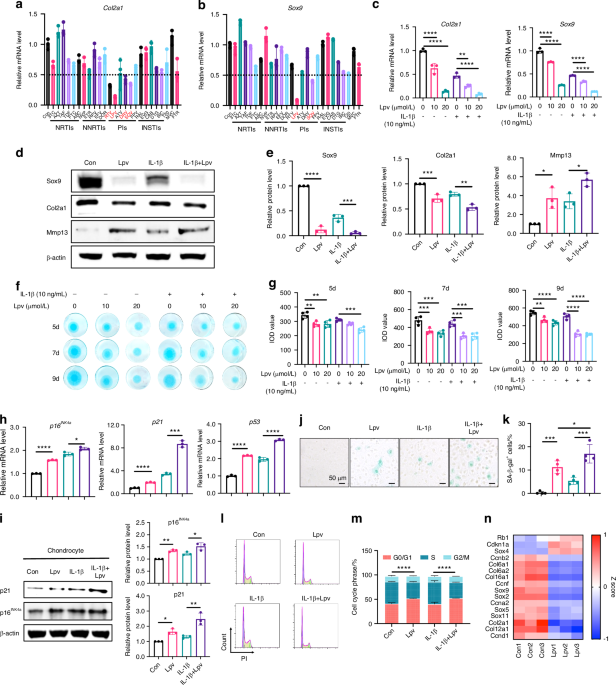
Loss of hepatic ApoE expression increases bone deposition in aged fracture callus. a Schematic diagram of the tibia fracture model in 24-month-old control (ApoEfl/fl, Ctrl) mice and liver-specific ApoE knockout (ApoEAlb, ΔApoE) mice. b ELISA was used to measure ApoE concentration in the plasma of mice (Ctrl, n = 10; ΔApoE, n = 15). c 21-day fracture calluses were assessed using micro-CT analysis to determine (d) total volume (TV), (e) bone volume (BV), and (f) bone ratio (BV/TV) (Ctrl, n = 9; ΔApoE, n = 13). g Safranin O/fast green/hematoxylin staining was used to stain the fracture calluses and (h) histomorphometric analysis was used to quantify the amount of bone within the fracture callus (Ctrl, n = 6; ΔApoE, n = 6). i Schematic diagram of tibial drill hole defect in 24-month-old control (ApoEfl/fl, Ctrl) mice and liver-specific ApoE knockout (ApoEAlb, ΔApoE) mice. j Healing was assessed 14 days after defect using micro-CT and (k) bone ratio (BV/TV) in the defect region was determined (Ctrl, n = 9; ΔApoE, n = 9). Data are presented as mean ± 95% confidence interval. *P < 0.05
Next, 24-month old mice underwent tibial fracture surgery and healing was measured 21 days post-injury using micro-CT and histological assessments. Micro-CT analysis demonstrated that while there was no difference in fracture callus size (total volume, TV), ΔApoE fracture calluses contained 50% more bone volume and had 50% higher bone ratio relative to Ctrl (Fig. 1c–f). Histological analysis corroborated these findings as Safranin O/fast green/hematoxylin staining of paraffin-embedded sections demonstrated a significant increase in bone tissue deposition in ΔApoE fracture calluses (Fig. 1g, h). Furthermore, osteoblast numbers were significantly increased in 21-day ΔApoE fracture calluses while cartilage template was decreased, and osteoclast activity was unchanged (Fig. S2).
To further assess differential bone healing, we used a tibial defect model. 24-month-old ΔApoE and Ctrl mice underwent tibial drillhole surgery and tissues were collected 14 days post-injury; healing was assessed by micro-CT (Fig. 1i). ΔApoE defects contained more bone tissue than did Ctrl defects (Fig. 1j, k) and osteoblast numbers were significantly increased while osteoclast activity was unchanged (Fig. S3).
Collectively, this work demonstrates that the primary source of circulating ApoE is the liver and that circulating ApoE impairs proper fracture healing with age.
ApoE impairs osteoblast differentiation while inhibiting the Wnt/β-catenin and Hippo pathwaysHere we show that loss of hepatic ApoE expression leads to increased osteoblast number and bone formation within the fracture callus. Thus, our next step was to determine ApoE’s effect on differentiation of aged bone marrow stromal cells (BMSCs) to osteoblasts. BMSCs from 24-month-old wildtype (C57BL/6 J) mice were cultured and differentiated to osteoblasts with vehicle or 100 ng/mL recombinant ApoE (rApoE) (Fig. 2a). Alkaline Phosphatase (Alk. Phos.) staining and Alizarin Red staining demonstrated that rApoE treatment decreased aged osteoblast differentiation and extracellular matrix mineralization (Fig. 2b). Furthermore, rApoE treatment was found to reduce expression of osteogenic marker genes Alp, Bsp, Col1α1, and Ocn in RT-PCR analysis (Fig. 2c–f). Interestingly, these findings were recapitulated when using cells from young, 4-month old mice. BMSCs from 4-month old mice were differentiated in osteogenic media containing vehicle or ApoE. As with aged cells, ApoE treatment inhibited osteoblastic differentiation of young cells (Fig. S4).
Fig. 2
ApoE impairs osteoblast differentiation and inhibits Wnt/β-catenin and Hippo signaling. a Schematic diagram of BMSC isolation, culture, subsequent osteogenic differentiation with vehicle or rApoE treatment and subsequent analysis. b BMSCs from 24-month-old C57BL/6 mice were cultured and differentiated in osteogenic media containing vehicle or rApoE. Wells were washed, fixed, and stained for Alkaline Phosphatase (Alk. Phos.) or mineral (Alizarin Red) to assess osteoblast differentiation and matrix mineralization, respectively. c–f Transcripts for osteogenic genes (Alp, Bsp, Col1, and Ocn) were measured using RT-PCR (n = 6). g Bulk RNA-seq was used to identify differentially expressed genes (DEGs) which were presented on a volcano plot using a log2(FC)-value of 2 and a P value of 0 as cut-offs for DEGs to compare vehicle-treated vs. rApoE-treated groups. h Downregulated DEGs were collected for Gene Ontology (GO) biological process analysis and (i) KEGG pathway analysis. j Western blot analysis of active-\(}\)-catenin and total \(}\)-catenin protein levels in vehicle-treated and rApoE-treated groups. k Immunofluorescence staining (active-\(}\)-catenin) of vehicle- and rApoE-treated osteogenic cultures. l, m Expression level of\(\,}\)-catenin target genes Axin2 and Cyclin D1 (n = 6). n Western blot analysis of active-Yap and total-Yap in vehicle-treated and rApoE-treated groups. o Immunofluorescence staining (active-Yap) of vehicle- and rApoE-treated osteogenic cultures. p–r Expression level Yap target genes Axl, Ctgf, and Cyr61 (n = 6). Data are presented as mean ± 95% confidence interval. *P < 0.05
We performed RNA sequencing (RNA-seq) on aged, ApoE-treated or untreated osteogenic cultures to identify the underlying mechanism of ApoE inhibition of osteoblast differentiation. Differentially expressed genes (DEGs) were depicted using a volcano plot (Fig. 2g) and biological functions of up- and down-regulated DEGs were assessed using gene ontology (GO) analysis. GO analysis indicated that the downregulated DEGs were mainly enriched in ossification, bone development, osteoblast differentiation and bone mineralization (Fig. 2h). We performed Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis of downregulated DEGs and enrichment analysis of KEGG revealed that the downregulated DEGs were mainly involved in the Wnt/β-catenin and the Hippo signaling pathways (Fig. 2i).
Gene set enrichment analysis (GSEA) demonstrated that the Wnt/β-catenin and the Hippo signaling pathways were significantly enriched (P < 0.01), a point that was further corroborated by heatmap visualization (Fig. S5). To validate modulation of these pathways, aged BMSCs were cultured in the absence or presence of ApoE and tested using biochemical techniques.
In the Wnt/β-catenin pathway, signaling is dependent on activation of β-catenin, its translocation to the nucleus, and subsequent expression of its transcriptional targets. Western blot analysis demonstrated that active β-catenin and total β-catenin protein levels decreased in response to ApoE treatment (Fig. 2j). Similarly, decreased active-β-catenin immunofluorescence staining was seen in cultures treated with ApoE (Fig. 2k) and Wnt/β-catenin pathway transcription targets, Axin2 and Cyclin D1, were down-regulated by ApoE treatment (Fig. 2l, m).
In the Hippo pathway, signaling is dependent on activation and subsequent nuclear translocation of the Yap/Taz protein complex into the nucleus to induce the TEAD-mediated gene transcription.12 Heatmap analysis of Hippo signaling pathway target genes indicated inhibition of this pathway (Fig. S5). Active-Yap and total-Yap protein levels decreased in response to ApoE treatment, as indicated by Western blot analysis and immunoblot staining (Fig. 2n, o). Transcriptional targets of the pathway (Axl, Cyr61, and Ctgf) were also found to be inhibited by ApoE treatment (Fig. 2p–r).
Collectively these findings indicate the involvement of the Wnt/β-catenin and/or Hippo pathways in ApoE-based inhibition of osteoblast differentiation.
Modulation of Wnt/β-catenin but not Hippo signaling is required for ApoE-based inhibition of osteoblast differentiationWe found both Wnt/β-catenin and Hippo signaling to be inhibited in concert with ApoE-based inhibition of osteoblast differentiation. To determine whether ApoE functions through the Wnt/β-catenin pathway, we cultured BMSCs from β-catenin-stabilized mice in which Cre-recombinase-based excision results in the production of a stabilized form of β-catenin protein. We chose this model over the loss-of-function β-catenin model because the loss-of-function model itself acts as a strong inhibitory model for osteoblast differentiation. Thus, interpretation of results assessing ApoE inhibitory activity layered on top of the loss-of-function inhibitory activity would be difficult. BMSC cultures from β-catenin stabilized mice were treated with adenovirus encoding green fluorescent protein (Ad-GFP, control) or adenovirus encoding a GFP-labelled Cre-recombinase (Ad-Cre) and were then differentiated in osteogenic media containing either vehicle or rApoE (Fig. 3a). Western blot analysis confirmed that ApoE-treatment lowered β-catenin levels in control samples but could not alter β-catenin levels in Ad-Cre cultures (Fig. 3b). Osteogenic differentiation was assessed by staining cultures, assessing type I collagen and osteocalcin levels, and quantifying osteogenic transcripts. ApoE treatment decreased staining in Ad-GFP control cultures but did not alter staining in Ad-Cre cultures for Alkaline Phosphatase (Fig. 3c) nor for Alizarin Red (Fig. 3d). Osteocalcin (OCN), a marker of osteoblast differentiation, was decreased by ApoE treatment in control cultures but did not change in Ad-Cre cultures (Fig. 3e). Finally, osteogenic transcripts (Alp, Bsp, Col1a1, Ocn) from rApoE-treated cultures were measured and related to osteogenic transcripts from vehicle-treated cultures. Ad-Cre-treated cultures were protected from ApoE-based inhibition while Ad-GFP were not (Fig. 3f–i).
Fig. 3
Modulation of the Wnt/\(}\)-catenin but not the Hippo pathway is required for ApoE’s inhibitory effect on osteoblast differentiation. a Schematic diagram of experimental design. BMSCs from mutant \(}\)-catenin loxP mice were cultured and treated with adenovirus carrying Cre-recombinase to generate \(}\)-catenin stabilized BMSCs (or carrying GFP as control). Cultures then underwent osteogenic differentiation in the presence of vehicle or rApoE. b Western blot analysis confirmed stabilization \(}\)-catenin in BMSCs upon treatment with adenovirus-Cre recombinase. c Cells were washed, fixed, and stained for Alkaline Phosphatase or (d) mineral (Alizarin Red) to assess osteoblast differentiation. e Western blot was used to assess the Col1α1 and OCN protein levels of osteogenic cultures. f–i Relative expression of osteogenic transcripts (Alp, Bsp, Col1a1, and Ocn) was measured in all culture conditions. Transcript expression in rApoE-treated cultures was related transcript expression of vehicle-treated cultures and plotted as Ad-GFP vs Ad-Cre. j Schematic diagram of experimental design. BMSCs from mutant Yap/Taz loxP mice were cultured and treated with adenovirus carrying Cre-recombinase to generate Yap/Taz knockout BMSCs (or carrying GFP as control). Cultures then underwent osteogenic differentiation in the presence of vehicle or rApoE. k Western blot analysis confirmed loss of Yap expression in BMSCs upon treatment with adenovirus-Cre recombinase. l Cells were washed, fixed, and stained for Alkaline Phosphatase or (m) mineral (Alizarin Red) to assess osteoblast differentiation. n Western blot was used to assess the Col1α1 and OCN protein levels of osteogenic cultures. o–r Relative expression of osteogenic transcripts (Alp, Bsp, Col1a1, and Ocn) was measured in all culture conditions. Transcript expression in rApoE-treated cultures was related transcript expression of vehicle-treated cultures and plotted as Ad-GFP vs Ad-Cre. Data are presented as mean ± 95% confidence interval. *P < 0.05
To determine whether ApoE functions through the Hippo pathway, we cultured BMSCs from Taz-floxed/Yap-floxed mice in which Cre-recombinase-based excision results in a loss of expression of the Yap and Taz proteins, muting transcriptional activity of the Hippo pathway. Similar to the tissue culture experiment above, cultures were treated with either adenovirus encoding green fluorescent protein (Ad-GFP, control) or adenovirus encoding a GFP-labelled Cre-recombinase (Ad-Cre) and were then differentiated in osteogenic media containing either vehicle or rApoE (Fig. 3j). Western blot analysis confirmed that ApoE-treatment significantly lowered Yap protein levels in control samples but could not alter Yap protein levels in Ad-Cre cultures (Fig. 3k). ApoE treatment decreased staining in Ad-GFP control cultures and likewise in Ad-Cre cultures for Alkaline Phosphatase (Fig. 3l) and for Alizarin Red (Fig. 3m). Furthermore, knockdown of Yap/Taz protein expression did not affect sensitivity to ApoE treatment. Both Ad-Cre and Ad-GFP treated cultures displayed similar levels of inhibition from ApoE as depicted by measurement of osteogenic transcripts (Alp, Bsp, Col1α1, Ocn) (Fig. 3n-q).
Collectively, these data indicate that ApoE-treatment of osteoblasts inhibits both the Wnt/β-catenin and Hippo pathways; however, ApoE’s osteoblast inhibitory activity functions through the Wnt/β-catenin pathway.
Lrp4 serves as the osteoblast cell-surface receptor for ApoEApoE cell-surface receptors are members of the low-density lipoprotein receptor (LDLR) family. The core members of the LDLR family expressed on osteoblasts include LDLR-related protein 1(LRP1), LRP2 (aka megalin), LRP4 (MEDF7), and LRP8 (aka ApoE receptor-2, ApoER2).13 To identify the osteoblast cell-surface receptor through which ApoE inhibits Wnt/β-catenin signaling and osteoblast differentiation, we used a slot blot apparatus to interrogate ApoE binding to Lrp2, Lrp4, Lrp8, and clusters 2, 3, and 4 of Lrp1. We found that Lrp4 bound ApoE with the highest abundance (Fig. 4a). Lrp4 is an evolutionarily conserved transmembrane protein in LDL receptor family. To further investigate whether Lrp4 from BMSCs can bind to ApoE, we overexpressed Flag-tagged Lrp4 in BMSCs and collected cell lysate to treat it with His-tagged ApoE. We then studied Lrp4-ApoE binding in BMSC lysates using co-immunoprecipitation. In lysates containing both Flag-Lrp4 and His-ApoE, immunoprecipitation of one protein also sequestered the other (Fig. 4b). Collectively these findings confirm ApoE binds Lrp4.
Fig. 4
Lrp4 is the osteoblast cell-surface receptor for ApoE. a Potential ApoE binding partners were blotted using slot blot and probed with ApoE. ApoE-binding was visualized using HRP-conjugated anti-ApoE antibody. b BMSCs were transfected with plasmid, inducing expression of Flag-tag Lrp4. Cells were then treated with His-labelled ApoE and lysates were immunoprecipitated using anti-Flag and/or anti-His antibodies to confirm ApoE-Lrp4 binding. c Schematic diagram depicting siRNA-based knockdown of Lrp4 in BMSCs from 24-month-old C57BL/6 mice. d Confirmation of Lrp4 gene knockdown in siRNA1-Lrp4 and siRNA2-Lrp4, but not siRNA-Ctrl. e–g Cells were washed, fixed, and stained for Alkaline Phosphatase (Alk. Phos.) or mineral (Alizarin Red) to assess osteoblast differentiation. h–k Relative expression of osteogenic transcripts (Alp, Bsp, Col1a1, and Ocn) was measured in all culture conditions. Transcript expression in rApoE-treated cultures was related transcript expression of vehicle-treated cultures and plotted as siRNA-Ctrl vs siRNA1-Lrp4 vs siRNA2-Lrp4. l Western blot analysis confirmed Lrp4 was required for ApoE-based modulation of β-catenin. Data are presented as mean ± 95% confidence interval. *P < 0.05
Next, we used siRNA to decrease the expression of Lrp4 in BMSCs to determine whether ApoE signals through Lrp4 to inhibit osteoblast differentiation (Fig. 4c). RT-PCR results demonstrated that our 2 different siRNAs, siRNA1-Lrp4 (siRNA165574-Lrp4) and siRNA2-Lrp4 (siRNA-94437-Lrp4), targeted Lrp4 and successfully inhibited Lrp4 expression by approximately 50% relative to the control siRNA-Ctrl (Fig. 4d). In siRNA-Ctrl cultures, ApoE treatment decreased the amount of Alkaline Phosphate and Alizarin Red staining. Importantly, knockdown of Lrp4 by siRNA1-Lrp4 or siRNA2-Lrp4 protected cultures from ApoE’s inhibitory effect as staining of cultures was unaffected by subsequent ApoE treatment (Fig. 4e–g). Furthermore, osteogenic transcripts (Alp, Bsp, Col1a1, and Ocn) were measured and normalized to siRNA-Ctrl. All four osteogenic transcripts were higher in siRNA1-Lrp4 and siRNA2-Lrp4 cultures than in siRNA-Ctrl (Fig. 4h–k). Finally, we tested the ability of ApoE treatment to modulate β-catenin levels in the absence of Lrp4. Active β-catenin levels were unchanged by ApoE treatment in siRNA1-Lrp4 and siRNA2-Lrp4 cultures while active β-catenin levels were decreased by ApoE-treatment in siRNA-Ctrl cultures, as demonstrated by Western blot analysis (Fig. 4l).
Collectively, these findings demonstrate that Lrp4 serves as the cell-surface receptor through which ApoE inhibits β-catenin signaling and osteoblast differentiation.
Human osteoblast differentiation is inhibited by ApoE though the Lrp4-Wnt/β-catenin axisThus far, we have used mouse models to demonstrate that circulating ApoE increases with age which impairs osteoblast differentiation by inhibiting the Wnt/β-catenin pathway through Lrp4-receptor signaling. As it does in mice, circulating ApoE increases with aging in human patients.10 We isolated human BMSCs (hBMSCs) from human bone marrow biopsies to determine whether ApoE can affect human osteoblast differentiation in a similar fashion as we see in mice. We used tissue culture techniques to assess 1) ApoE-dependent changes in osteoblast differentiation, 2) ApoE-dependent changes in Wnt/β-catenin signaling, and 3) the reliance on Lrp4 for ApoE-dependent changes (Fig. 5a).
Fig. 5
Human osteoblast differentiation is inhibited by ApoE through Lrp4-Wnt/β-catenin signaling. a Schematic diagram, human BMSCs were isolated from bone marrow, cultured, and differentiated in osteogenic media with vehicle or rApoE. b Cells were washed, fixed, and stained for Alkaline Phosphatase or mineral (Alizarin Red) to assess osteoblast differentiation. c–f Osteogenic transcripts (Alp, Bsp, Col1, and Ocn) were measured using RT-PCR (n = 6). g Western blot analysis of active-\(}\)-catenin and total \(}\)-catenin protein levels in vehicle-treated and rApoE-treated groups. h, i Expression level of\(\,}\)-catenin target genes Axin2 and Cyclin D1 (n = 6). j Confirmation of Lrp4 gene knockdown in siRNA1-Lrp4. k, l Cells were washed, fixed, and stained for Alkaline Phosphatase (Alk. Phos.) or mineral (Alizarin Red) to assess osteoblast differentiation. m–p Relative expression of osteogenic transcripts (Alp, Bsp, Col1a1, and Ocn) was measured in all culture conditions. Transcript expression in rApoE-treated cultures was related transcript expression of vehicle-treated cultures and plotted as siRNA-Ctrl vs siRNA-Lrp4. Data are presented as mean ± 95% confidence interval. *P < 0.05
First, hBMSCs were differentiated in osteogenic media in the presence of vehicle or rApoE. Alkaline Phosphate and Alizarin Red staining demonstrated that ApoE treatment decreased osteogenic activity and decreased matrix mineralization in these cells (Fig. 5b). Likewise, RT-PCR revealed that the expression level of osteoblast differentiation markers (Alp, Bsp, Col1a1, and Ocn) was significantly reduced in response to ApoE-treatment (Fig. 5c–f).
Next, we assessed changes in Wnt/β-catenin signaling in response to ApoE treatment. Western blot analysis of protein lysates demonstrated that ApoE treatment of hBMSCs decreased active and total β-catenin levels in osteogenic cultures (Fig. 5g). Furthermore, RT-PCR analysis of transcripts showed that ApoE treatment significantly decreased expression levels of β-catenin target genes Axin2 and Cyclin D1 (Fig. 5h, i).
Lastly, we used our siRNA system to investigate the role of Lrp4 in ApoE-induced inhibition of osteoblast differentiation. hBMSCs were cultured and transfected with either siRNA-Ctrl, siRNA1-Lrp4, siRNA2-Lrp4, or siRNA-3-Lrp4. siRNA1-Lrp4 successfully lowered Lrp4 levels (Fig. 5j). Fresh hBMSCs were then cultured and transfected with either siRNA-Ctrl or siRNA1-Lrp4. Cultures were maintained in osteogenic media with vehicle or rApoE. ApoE treatment had no effect on siRNA1-Lrp4-treated cultures but decreased Alkaline Phosphatase and Alizarin Red staining of siRNA-Ctrl-treated cultures (Fig. 5k, l). Next, hBMSCs treated with siRNA-Ctrl or siRNA1-Lrp4 were cultured in osteogenic media containing ApoE. The level of osteogenic transcripts was compared between the two groups to assess the impact of Lrp4 knockdown in ApoE-based inhibition of osteoblast differentiation. In siRNA1-Lrp4 cultures, knockdown of Lrp4 rendered the cells insensitive to ApoE treatment (Fig. 5m–p).
Collectively, these data demonstrate that similar to the mechanism we identified in mouse models, ApoE inhibits human BMSC-to-osteoblast differentiation by binding to Lrp4 and inhibiting Wnt/β-catenin signaling.
Neutralizing circulating ApoE improves bone fracture healing in aged miceNext, we tested the ability of an ApoE-neutralizing antibody to transiently lower circulating ApoE and counteract the negative effects on bone regeneration. To this end, we tested the ability of an ApoE-neutralizing antibody (NAb, HJ6.3) to improve aged fracture healing (Fig. 6a).
Fig. 6
ApoE-neutralizing antibody improves aged bone healing. a Schematic diagram of fractured 24-month-old mice treated with ApoE-neutralizing antibody and assessed for fracture healing. b Mice were injected with IgG or NAb 3 days post injury and blood was collected 7 days post-injection and assessed for ApoE concentration (n = 5). c Micro-CT analysis of 21-day fracture calluses was used to determine (d) total volume (TV), (e) bone volume (BV), (f) bone deposition (BV/TV), and (g) tissue mineral density (n = 8). h Safranin O/fast green staining was used to stain the fracture calluses and (i) histomorphometric analysis was used to quantify the amount of bone within fracture calluses (n = 6). Data are presented as mean ± 95% confidence interval. *P < 0.05
24-month-old mice were treated with IgG control or NAb (10 mg/kg, IP). Plasma collected 7 days post-injection showed a 75% decrease in circulating ApoE concentrations (Fig. 6b). Liver panels and lipid levels were performed using HPLC/MS: total cholesterol, HDL, LDL, triglycerides, albumin, AST, and ALT were all measured in samples from IgG control and NAb-treated mice. Cholesterol, HDL, and LDL were all unchanged, indicated no significant changes in plasma lipid levels (Fig. S6). ALT was also unchanged however; triglycerides were slightly increased in NAb samples while albumin and AST were slightly decreased in NAb samples (Fig. S6). Therefore, while there were slight changes in triglycerides, albumin, and AST, NAb treatment was successful in lowering circulating ApoE levels.
Next, 24-month-old mice underwent tibial fracture surgery and were treated with NAb or IgG 3 days after injury. Fracture calluses were collected 21 days post-fracture and assessed using micro-CT and histological techniques). Micro-CT analysis of fracture calluses demonstrated that NAb treatment did not change the total volume; however, bone volume, bone ratio, and tissue mineral density were significantly increased (Fig. 6c–g). Moreover, histomorphometry of Safranin O/fast green/hematoxylin stained sections revealed that fracture callus of the NAb-treated group contained approximately 50% more bone tissue than did the IgG-treated control group (Fig. 6h, i). Furthermore, cartilage formation and osteoclast activity was unaffected 14-day post fracture (Fig. S7) however, osteoblast numbers were significantly increased in 21-day NAb-treated fracture calluses while cartilage template was decreased, and osteoclast activity was unchanged (Fig. S8).
Importantly, circulating ApoE levels and fracture healing was unaffected by NAb treatment of young mice (Fig. S9). Collectively, these data demonstrate that treatment with ApoE-neutralizing antibody lowers circulating ApoE levels in aged mice which significantly improves bone fracture healing.
Comments (0)