
Remember me
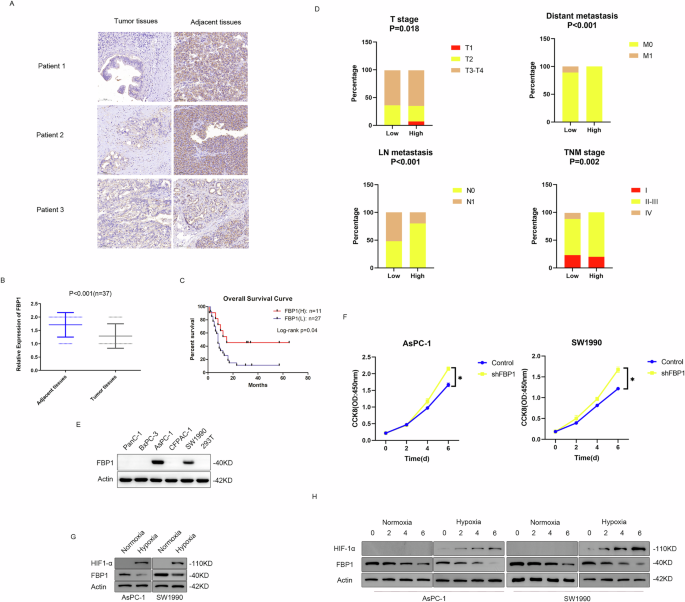
First, we examined the expression of FBP1 in PDAC samples to obtain a comprehensive understanding of FBP1. An evaluation of three pairs of representative images revealed that FBP1 expression in the tumour tissues was lower than that in the matched adjacent tissues (Fig. 1A). Statistical analysis revealed that FBP1 expression was lower in PDAC tissues than in adjacent normal pancreatic tissues (P < 0.001) (Fig. 1B). Next, we demonstrated that high FBP1 expression in tumour tissues usually predicted a better prognosis in patients (P = 0.04) (Fig. 1C). Conversely, low FBP1 expression in PDAC tissues was negatively correlated with advanced T stage (P = 0.018), lymph node metastasis (P < 0.001), distant metastasis (P < 0.001) and late AJCC TNM stage (P = 0.002), suggesting that FBP1 downregulation contributed to PDAC tumorigenesis and progression in these 38 clinical cases (Fig. 1D). Therefore, additional pathways leading to FBP1 protein degradation need to be discovered. FBP1 protein expression levels were detected in pancreatic cancer lines, and we found that FBP1 was expressed in the AsPC-1 and SW1990 cell lines. Therefore, these two cell lines were used for further investigation (Fig. 1E). FBP1 knockdown in the sPC-1 and SW1990 cell lines accelerated cell growth, indicating that FBP1 is a tumour suppressor in these two cell lines (Fig. 1F). Because PDAC tissue is usually reported to be hypoxic, we investigated whether there is a relationship between hypoxia and FBP1. To validate the effect of hypoxia on FBP1, we exposed the pancreatic cancer cell lines AsPC-1 and SW1990 to hypoxia and found that FBP1 expression was downregulated (Fig. 1G). A cycloheximide (CHX) chase assay was subsequently performed, and the results revealed that the protein stability of FBP1 decreased under hypoxic conditions (Fig. 1H and Supplementary Fig. 1).
Fig. 1: Downregulation of FBP1 expression predicts a poor prognosis in patients with pancreatic cancer, and O-GlcNAcylation regulates FBP1 expression under hypoxia.
A Representative IHC staining of 38 pairs of pancreatic tumour and normal adjacent tissues. Scale bar as shown. B Statistical analysis of FBP1 expression from the IHC results. We defined high expression as two points and low expression as 1 point. C Kaplan–Meier overall survival curves for all 38 patients with pancreatic cancer stratified by high and low FBP1 expression. D Correlations of FBP1 expression with T stage, lymph node metastasis, distant metastasis and AJCC TNM stage in clinical patients with pancreatic cancer according to the χ2 test. E Screening of FBP1 expression in pancreatic cancer cell lines. F Downregulation of FBP1 accelerates tumour growth in AsPC-1 and SW1990 cells. G FBP1 is repressed under hypoxia in AsPC-1 and SW1990 cells. H A hypoxic environment triggered the degradation of FBP1 in AsPC-1 and SW1990 cells.
OGT binds to FBP1 and promotes FBP1 O-GlcNAcylation at serine 47A previous study reported that hypoxia can induce hyper-O-GlcNAcylation. As the exposure time to the hypoxic environment increased, the O-GlcNAcylation level gradually increased, and FBP1 expression gradually decreased (Fig. 2A). Then, we used the siOGT plasmid and OSMI-1 (an OGT inhibitor) to treat AsPC-1 and SW1990 cells that had been subjected to hypoxia for 6 h. Treatment with either siOGT or OSMI-1 dramatically reduced O-GlcNAcylation. Moreover, FBP1 expression was elevated (Fig. 2B). Collectively, these findings suggest that O-GlcNAcylation signalling might participate in hypoxia-induced FBP1 degradation.
Fig. 2: OGT interacts with and O-GlcNAcylates FBP1 at the serine 47 residue.
A Time course of FBP1 expression and hypoxia exposure time. B Treatment with siOGT or OSMI-1 reversed the degradation of FBP1 in AsPC-1 and SW1990 cells under hypoxia for 6 h. C A co-IP assay was performed to validate the interaction between exogenous FBP1/FBP1 (K908A) and OGT plasmids in 293 T cells. D A co-IP assay was performed to validate the interaction between endogenous FBP1 and OGT in AsPC-1 and SW1990 cells. E A PLA was performed to validate the interaction between endogenous FBP1 and OGT in AsPC-1 cells. The PLA signal was detected by fluorescence microscopy (red arrows). Scale bar as shown. F Four FBP1 truncations were constructed according to their structural domains. A co-IP assay was performed to determine the interacting domain. G Mass spectrometry results of the FBP1 serine 47 residue. H A co-IP assay between O-GlcNAc and the FBP1-WT/mutant was performed to validate the potential O-GlcNAcylation site. Endogenous FBP1 was knocked down and then rescued by Flag-tagged FBP1-WT, S47A, S67A, and S233A plasmids in AsPC-1 and SW1990 cells.
We hypothesised that there is a relationship between O-GlcNAcylation and FBP1. To test this hypothesis, we performed an exogenous IP assay, and the results revealed that FBP1 can interact with OGT in the 293T cell line. 293T cells were also transfected with the OGT mutant K908A (with loss of O-GlcNAcylation activity), and this mutation was found to disrupt FBP1 O-GlcNAcylation and increase FBP1 protein expression. However, the interaction between the two proteins was not disrupted (Fig. 2C). Next, endogenous IP assays were performed in AsPC-1 and SW1990 cells, and the results indicated that endogenous FBP1 could be O-GlcNAcylated via interaction with endogenous OGT (Fig. 2D). A proximity ligation assay (PLA) was performed to further validate the interaction between OGT and FBP1, and red fluorescent puncta were detected mainly in the cytoplasm (Fig. 2E). We next sought to determine which domain of FBP1 interacts with OGT. To this end, four truncations of FBP1 were constructed according to its functional domains. OGT mainly interacted with the Δ1 domain of FBP1 (exons 1 and 2) (Fig. 2F).
To identify the O-GlcNAcylation site(s) on FBP1, we performed protein purification and mass spectrometry analysis, and two potential O-GlcNAcylation sites (S47 and S211) on FBP1 were revealed by LC–MS/MS (Fig. 2G and Supplementary Fig. 2). A co-IP assay was subsequently performed with the wild-type (WT) FBP1 protein and three FBP1 site mutants (S47A, S211A and S67A (negative control); serine-to-alanine mutants), and the results revealed that the S47A mutation, but not the S211A or S67A mutation, significantly reduced the O-GlcNAcylation level of FBP1 (Fig. 2H). Therefore, we chose the S47A mutant for further investigation.
FBP1 O-GlcNAcylation at S47 can promote its degradationTo determine whether the S47A mutation interferes with FBP1 protein degradation, we constructed two stably transfected AsPC-1 and SW1990 cell lines (FBP1-WT and FBP1-S47A). We subsequently downregulated O-GlcNAcylation in these two cell lines by transfecting two siOGT plasmids. The western blot results revealed that the downregulation of OGT in the FBP1-WT cell line increased the protein expression of FBP1. However, FBP1 expression did not change in the FBP1-S47A cell line under the same conditions (Fig. 3A). In contrast, OGT downregulation did not change the mRNA level of FBP1 in either cell line, suggesting the possibility of posttranslational regulation (Fig. 3B). To determine whether O-GlcNAcylation can affect the protein stability of FBP1, we performed a CHX chase assay in AsPC-1 and SW1990 cells, and the results revealed that OGT knockdown increased the protein stability of FBP1-WT. However, cells transfected with FBP1-S47A did not exhibit this effect (Fig. 3C and Supplementary Fig. 3). Similarly, forced O-GlcNAcylation by transfection of the OGT plasmid but not the OGT K908A mutant plasmid resulted in decreased FBP1 expression, although this effect was not observed in cells transfected with FBP1-S47A (Fig. 3D), suggesting that the OGT-mediated change in FBP1 protein expression was dependent on the catalytic function of OGT. In addition, OGT overexpression did not affect the FBP1 mRNA expression in either cell line (Fig. 3E). We also performed a CHX chase assay, and the results revealed that OGT overexpression decreased the protein stability of FBP1-WT. However, cells transfected with FBP1-S47A did not exhibit this effect (Fig. 3F and Supplementary Fig. 4). Next, we stimulated these two cell lines (FBP1-WT and FBP1-S47A) in a hypoxic environment and found that hypoxia dramatically downregulated FBP1 expression in the FBP1-WT cells compared with the FBP1-S47A cells (Fig. 3G). A CHX chase assay was performed, and the results revealed that a hypoxic environment decreased the protein stability of FBP1-WT to a greater extent (Fig. 3H and Supplementary Fig. 5). Together, these results indicate that FBP1 O-GlcNAcylation at serine 47 can promote FBP1 protein stability in a posttranslational manner.
Fig. 3: FBP1 O-GlcNAcylation at the serine 47 residue promotes its degradation in a hypoxic environment.
A Silencing OGT with siRNAs regulates FBP1-WT/S47A protein expression levels in AsPC-1 and SW1990 cells. B Silencing OGT with siRNAs regulates FBP1-WT/S47A mRNA expression levels in AsPC-1 and SW1990 cells. C CHX assays were performed to determine the stability of the FBP1-WT/S47A proteins in OGT-knockdown AsPC-1 and SW1990 cells. D Hyper-O-GlcNAcylation by OGT or an OGT mutant (K908A) regulates FBP1-WT/S47A protein expression levels in AsPC-1 and SW1990 cells. E Hyper-O-GlcNAcylation by OGT or an OGT mutant (K908A) regulates FBP1-WT/S47A mRNA expression levels in AsPC-1 and SW1990 cells. F CHX assays were performed to determine the stability of the FBP1-WT/S47A proteins under hyper-O-GlcNAcylation by OGT in AsPC-1 and SW1990 cells. G Changes in FBP1-WT/S47A protein expression levels in AsPC-1 and SW1990 cells under hypoxic conditions. H CHX assays were performed to determine the stability of the FBP1-WT/S47A proteins in a hypoxic environment in AsPC-1 and SW1990 cells.
FBP1 O-GlcNAcylation at S47 influences the expression of its downstream target genes and promotes cancer progressionBecause FBP1 can inhibit the onset of the Warburg effect by reducing the expression of canonical HIF-1α target genes (PDK1, LDHA, GLUT1 and VEGF), we performed qRT‒PCR to confirm whether FBP1 O-GlcNAcylation at serine 47 can affect the expression of downstream target genes under hypoxic conditions. Compared with the FBP1-WT group, the FBP1-S47A group presented decreased expression of target genes (PDK1, LDHA, GLUT1 and VEGF) induced by FBP1 overexpression. Furthermore, OGT knockdown via shRNA decreased the expression levels of these target genes in the AsPC-1 and SW1990 cell lines, but no significant difference was found between the FBP1-WT and FBP1-S47A groups (Fig. 4A). Chromatin immunoprecipitation (ChIP) assays of the AsPC-1 and SW1990 cell lines revealed that FBP1-WT was enriched at the hypoxia response elements (HREs) of the PDK1, LDHA, GLUT1 and VEGF promoters but not at the promoter of RPL13A, which is nonresponsive to hypoxia. The FBP1-S47A mutation or OGT knockdown increased FBP1 enrichment at the HREs of the PDK1, LDHA, GLUT1 and VEGF promoters, which may enhance the inhibitory effect of FBP1 on the expression of its target genes (Fig. 4B).
Fig. 4: FBP1 O-GlcNAcylation at the serine 47 residue regulates its downstream target genes and promotes tumour progression.
A RT‒qPCR results showing the expression levels of downstream target genes in AsPC-1 and SW1990 (FBP1-WT/S47A) cells with or without OGT knockdown. B ChIP‒qPCR results showing FBP1 occupancy at the promoters of downstream target genes in AsPC-1 and SW1990 (FBP1-WT/S47A) cells with or without OGT knockdown. C Glucose uptake assays were performed in AsPC-1 and SW1990 (FBP1-WT/S47A) cells with or without OGT knockdown. D Lactate secretion assays were performed in AsPC-1 and SW1990 (FBP1-WT/S47A) cells with or without OGT knockdown. E MTT assays were performed to measure the growth of AsPC-1 and SW1990 (FBP1-WT/S47A) cells with or without OGT knockdown. F Colony formation assays were performed in AsPC-1 and SW1990 (FBP1-WT/S47A) cells with or without OGT knockdown, and the statistical results were calculated. G Xenograft experiments in nude mice were performed in AsPC-1 and SW1990 (FBP1-WT/S47A) cells with or without OGT knockdown, and the statistical results (i.e., tumour weight and volume) were analysed.
Next, we measured glucose metabolism in pancreatic cancer cell lines. The FBP1-S47A mutation can reduce glucose uptake and lactate production. OGT knockdown also reduced glucose uptake and lactate production in the FBP1-WT group but had no effect on the FBP1-S47A mutant group (Fig. 4C, D), which was consistent with the results described above for the expression of FBP1 target genes. Compared with WT FBP1, the expression of the FBP1-S47A mutant suppressed AsPC-1 and SW1990 cell growth (Fig. 4E). Indeed, OGT knockdown promoted colony formation by FBP1-WT cells but did not further inhibit colony formation by FBP1-S47A mutant cells (Fig. 4F). Consistent with the in vitro results, OGT knockdown significantly reduced xenograft tumour growth (i.e. tumour volume and weight) in mice injected with FBP1-WT cells but had no significant effect on tumour growth in mice injected with the FBP1-S47A mutant cells, suggesting that the suppressive effect may be largely dependent on FBP1 O-GlcNAcylation at serine 47 in vivo (Fig. 4G).
FBP1 O-GlcNAcylation at S47 can promote its K48 polyubiquitinationFBP1 has been shown to be O-GlcNAcylated at serine 47 to promote its degradation and tumour progression under hypoxic conditions. Given that ubiquitination is usually involved in protein degradation, we sought to determine whether ubiquitination could play a role in the O-GlcNAcylation-mediated degradation of FBP1. First, we transfected plasmids expressing Ub and the E3 ligase TRIM28 into pancreatic cancer cells to increase the activity of the ubiquitination system, and we found that OGT (but not OGT 908 A) expression increased FBP1 polyubiquitination and that this effect can be abolished by S47 mutation (Fig. 5A). Because hypoxia can induce hyper-O-GlcNAcylation, we exposed cancer cell lines to hypoxia and found that hypoxia promoted both the polyubiquitination and the degradation of FBP1 in the FBP1-WT cell group (Fig. 5B). Next, we found that the E3 ligase TRIM28 catalysed FBP1 polyubiquitination in a K48-dependent manner and that O-GlcNAcylation at S47 promoted this process (Fig. 5C). However, the K63-dependent polyubiquitination process of FB1 was very weak in this system, and O-GlcNAcylation at S47 did not affect it (Fig. 5D). To determine the potential effect of TRIM28-mediated K48 polyubiquitination, TRIM28 was knocked down, and the results revealed that the K48 polyubiquitination level dramatically decreased in both the FBP1-WT and FBP1-S47 groups without affecting the O-GlcNAcylation level of FBP1 (Fig. 5E). We then replaced OGT overexpression with a hypoxic environment, and the results showed that silencing TRIM28 could partially reverse FBP1 O-GlcNAcylation-mediated degradation (Fig. 5F).
Fig. 5: FBP1 O-GlcNAcylation at the serine 47 residue promotes its TRIM28-mediated K48 polyubiquitination.
A Cotransfection of OGT and an OGT mutant (K908A) revealed that OGT facilitates TRIM28-mediated FBP1 polyubiquitination in AsPC-1 and SW1990 wild-type cells. B Hypoxia facilitates TRIM28-mediated FBP1 polyubiquitination in AsPC-1 and SW1990 wild-type cells. C OGT facilitates TRIM28-mediated FBP1 K48-linked polyubiquitination in AsPC-1 and SW1990 wild-type cells. D OGT fails to affect TRIM28-mediated FBP1 K63-linked polyubiquitination in AsPC-1 and SW1990 wild-type cells. E Silencing TRIM28 abolishes OGT-mediated protein degradation in AsPC-1 and SW1990 wild-type cells. F Silencing TRIM28 abolishes hypoxia-mediated protein degradation in AsPC-1 and SW1990 wild-type cells.
FBP1 polyubiquitination at K51 is dependent on S47 O-GlcNAcylationTo identify potential polyubiquitination sites in the FBP1 protein, the gel band containing ubiquitinated FBP1 was excised and analysed by LC–MS/MS. As shown in Supplementary Figs. 5A, 6, five potential ubiquitination sites in FBP1 were identified. We subsequently constructed five lysine (K)-to-arginine (R) mutants (K51R, K110R, K143R, K151R and K232R) and performed a co-IP assay to determine the functional sites. Mutation of only the K51R site attenuated FBP1 K48 polyubiquitination (Fig. 6B). The “SAVRK” sequence is highly conserved in many species (Fig. 6C). We subsequently performed a CHX chase assay to determine whether this mutant (K51R) can affect FBP1 protein stability, and the results revealed that, compared with FBP1-WT, FBP1-K51R was less prone to degradation (Fig. 6D and Supplementary Fig. 7). We next sought to determine whether there is a relationship between FBP1 O-GlcNAcylation at S47 and FBP1 polyubiquitination at K5. Interestingly, compared with the FBP1-WT group, OGT transfection changed the O-GlcNAcylation level in the FBP1-K51R group but not in the FBP1-S47A group. However, the polyubiquitination level increased in the FBP1-WT group but changed slightly in the FBP1-K51R and FBP1-S47A groups (Fig. 6E). In addition, hypoxia led to increased O-GlcNAcylation in the FBP1-WT and FBP1-K51R groups but not in the FBP1-S47A group, and this increase was partially reversed by siOGT transfection. Moreover, hypoxia resulted in greater K48 polyubiquitination in the FBP1-WT group than in the other groups, and this effect was inhibited in both the FBP1-S47A and FBP1-K51R groups (Fig. 6F). Collectively, these data suggest that O-GlcNAcylation at serine 47 might be a prerequisite for FBP1 polyubiquitination at lysine 51 in the context of hypoxia-induced protein degradation.
Fig. 6: FBP1 O-GlcNAcylation at the serine 47 residue promotes its K51 polyubiquitination.
A Mass spectrometry results of the FBP1 serine 47 residue. B Co-IP between Ub and the FBP1-WT/lysine mutants was performed to validate the potential polyubiquitination site. C Conserved domains in the FBP1 protein in many species. D CHX assays were performed to determine the stability of the FBP1-WT/K51R proteins in AsPC-1 and SW1990 cells. E Cotransfection of OGT and Ub plasmids revealed that blocking FBP1 O-GlcNAcylation at the serine 47 residue can reverse its K51 polyubiquitination in AsPC-1 and SW1990 cells. F Blocking the O-GlcNAcylation of FBP1 at the serine 47 residue can reverse the K51 polyubiquitination of FBP1 in AsPC-1 and SW1990 cells induced by a hypoxic environment.
FBP1 polyubiquitination at K51 influences the expression of its downstream target genes and promotes cancer progressionBecause FBP1 K51 polyubiquitination is the downstream effect of its O-GlcNAcylation, we measured the expression levels of canonical HIF-1α target genes (PDK1, LDHA, GLUT1 and VEGF) in the FBP1-WT and FBP1-K51R groups. Mutation of the K51 site increased the expression levels of these target genes under hypoxic conditions, and these increases were partially reversed by OGT knockdown in pancreatic cancer cells (Fig. 7A). Finally, to further confirm that the K51 polyubiquitination of FBP1 is downstream of its O-GlcNAcylation during pancreatic cancer progression, we repeated the functional experiments with FBP1-WT and FBP1-K51R cells. Consistent with the previous results, OGT knockdown in the FBP1-WT cells reduced glucose uptake and lactate production but had no such effect on the FBP1-K51R mutant cells (Fig. 7B, C). Compared with the expression of FBP1-WT, the expression of the FBP1-K5R mutant suppressed AsPC-1 and SW1990 cell growth (Fig. 7D). OGT knockdown also promoted colony formation by FBP1-WT cells but did not further inhibit colony formation by FBP1-K51R mutant cells (Fig. 7E). Consistent with the in vitro results, OGT knockdown significantly reduced xenograft tumour growth (i.e., tumour volume and weight) in mice injected with FBP1-WT cells but had no significant effect on tumour growth in mice injected with FBP1-K51R mutant cells, suggesting that the effect of K51 polyubiquitination may be dependent mainly on O-GlcNAcylation activity in vivo (Fig. 7F). In summary, we showed that hypoxia results in the deactivation and degradation of FBP1 in pancreatic cancer cells both in vivo and in vitro via the serine 47 O-GlcNAcylation-lysine 51 polyubiquitination axis (Fig. 7G).
Fig. 7: FBP1 K51 polyubiquitination regulates its downstream target genes, and the O-GlcNAcylation/polyubiquitination axis promotes tumour progression.
A RT‒qPCR results showing the expression levels of downstream target genes in AsPC-1 and SW1990 (FBP1-WT/K51R) cells with or without OGT knockdown. B Glucose uptake assays were performed in AsPC-1 and SW1990 (FBP1-WT/K51R) cells with or without OGT knockdown. C Lactate secretion assays were performed in AsPC-1 and SW1990 (FBP1-WT/K51R) cells with or without OGT knockdown. D MTT assays were performed to measure the growth of AsPC-1 and SW1990 (FBP1-WT/K51R) cells with or without OGT knockdown. E Colony formation assays were performed in AsPC-1 and SW1990 (FBP1-WT/K51R) cells with or without OGT knockdown, and the statistical results were calculated. F Xenograft experiments in nude mice were performed in AsPC-1 and SW1990 (FBP1-WT/K51R) cells with or without OGT knockdown, and the statistical results (i.e. tumour weight and volume) were analysed. G Schematic model of how the FBP1 O-GlcNAcylation/polyubiquitination axis functions in pancreatic cancer cells in a hypoxic environment.
Comments (0)