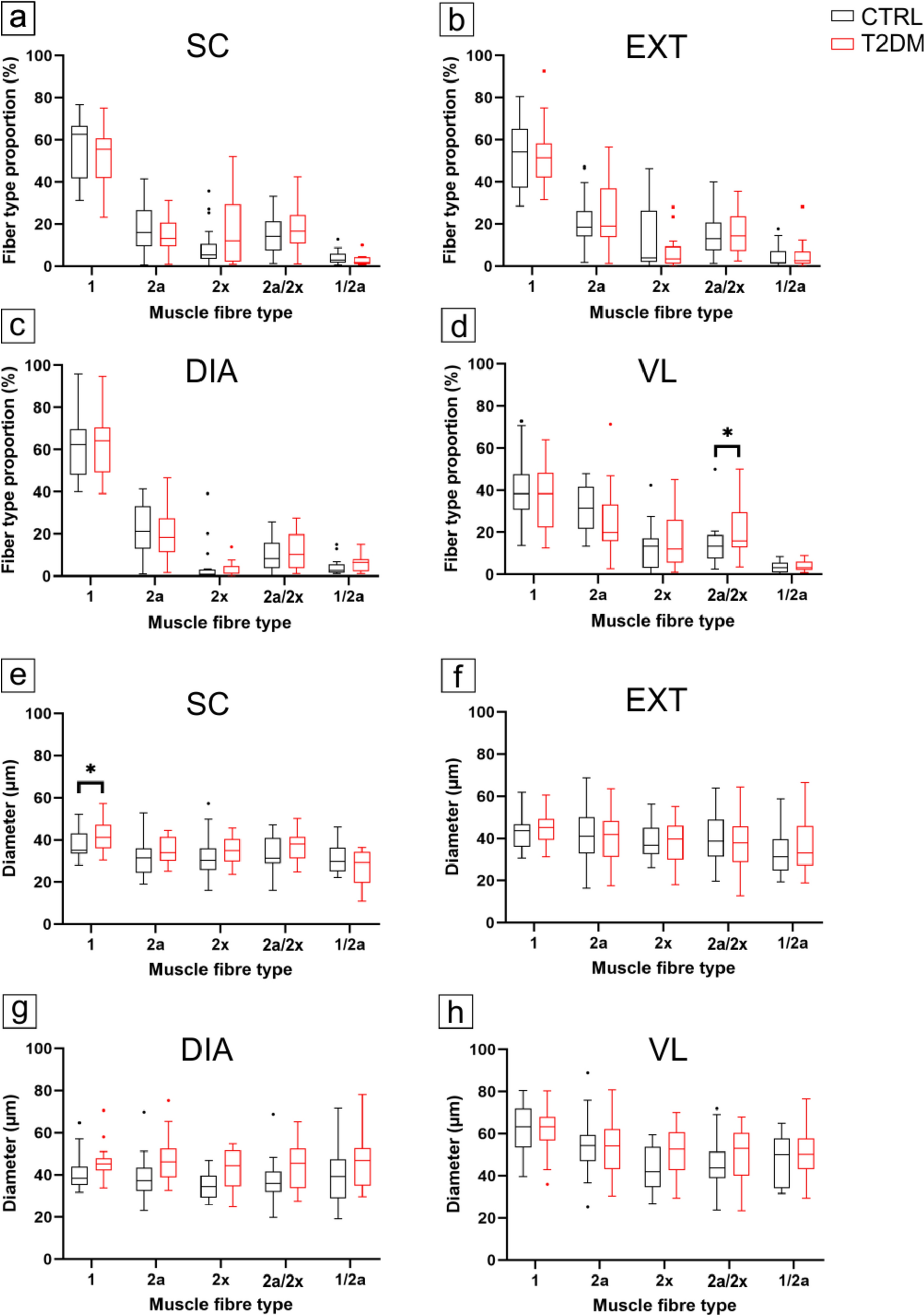
This study analysed four functionally distinct skeletal muscles, SC, DIA, EXT and VL, obtained from the same older men with and without T2DM, using MyHC-based fibre typing, Sudan Black B IMCL staining and 3D confocal capillary morphometry. The T2DM group had HbA1c values generally within commonly targeted ranges and no documented advanced diabetic complications, supporting interpretation of the findings as early or moderate structural differences associated with diabetes and adiposity rather than overt advanced myopathy.
Overall, group differences were modest and muscle-specific, superimposed on a preserved hierarchy across muscles. Fibre-type composition and mean fibre diameter were largely maintained; IMCL was higher in SC and EXT; oxidative fibres (types 1 and 2a) contained more IMCL than type 2x fibres in both groups; and LVf was selectively reduced in the DIA, while other capillary indices were largely similar between groups. Among capillary endpoints, the DIA showed the clearest group difference, a selective reduction in LVf. SC showed the clearest fibre-size difference (larger type 1 fibres), EXT exhibited a small change in anisotropy and VL showed a modest increase in 2a/2x hybrids together with BMI-associated IMCL variation. Together, the pattern of results is consistent with a dominant influence of muscle-specific functional demands and oxidative phenotype, and with adiposity as an important covariate, rather than a uniform diabetes effect across the musculature, consistent with previous morphological and microvascular studies in diabetic and obese muscle (Umek et al. 2019).
The preservation of fibre-type composition in the postural (SC) and respiratory (DIA, EXT) muscles, and the modest increase in 2a/2x hybrids in VL, contrasts with reports of a more pronounced shift toward faster, more glycolytic phenotypes in some locomotor muscles in obesity and T2DM (Park et al. 2009; Andreassen et al. 2014). The larger SC type 1 fibres in T2DM may reflect chronic loading associated with higher body mass and sustained postural activity, although causal inference is limited by the cross-sectional, post-mortem design. Similar preservation or mild hypertrophy of fibres in frequently recruited muscles has been reported in models of obesity-induced insulin resistance, where increased mechanical load supports fibre growth despite systemic metabolic stress (Ato et al. 2019; Umek et al. 2021a).
Continuously active postural and respiratory muscles are exposed to sustained mechanical and metabolic demand, which can favour anabolic signalling and mitochondrial maintenance and may oppose atrophy programmes mediated by FoxO transcription factors (Sandri et al. 2006; Ogasawara et al. 2013). Conversely, intermittently recruited muscles, such as VL, may be more sensitive to reductions in habitual activity, particularly in older individuals. The fibre-type-specific age associations observed in DIA, EXT and VL should be interpreted as cohort-specific and may reflect selective survival of larger fibres, unmeasured activity differences or other confounding, rather than a general ageing signature. The minimal influence of BMI and HbA1c on fibre size suggest that, in this cohort of older men, muscle-specific factors, likely related to regional workload and oxygen demand, outweigh systemic factors such as BMI in determining fibre size (Frontera et al. 2000; Cameron et al. 2023). Collectively, these findings indicate that diabetes does not uniformly impair myofibre morphology; continuously active muscles, such as the postural SC, retain or even modestly increase their fibre calibre, which may reflect functional adaptation to sustained contractile and metabolic demands rather than pathological hypertrophy.
IMCL was modestly higher in T2DM, most clearly in SC and EXT, indicating that habitual postural or respiratory activity does not preclude greater lipid storage when adiposity is higher. This is consistent with evidence that IMCL content reflects both oxidative phenotype and lipid availability, and that IMCL per se is not a direct surrogate of insulin sensitivity without information on lipid species, subcellular localisation and turnover (Krssak et al. 1999; Goodpaster et al. 2001; Dubé et al. 2008; Coen and Goodpaster 2012). In the present study, IMCL was quantified as the percentage of fibre area occupied by Sudan Black B-positive lipid droplets, so this readout reflects bulk intracellular lipid staining on two-dimensional sections rather than lipid composition or subcellular droplet organisation. It does not distinguish lipid species, droplet size or subsarcolemmal from intermyofibrillar localisation. This limits mechanistic inference because ultrastructural studies in human skeletal muscle indicate that insulin resistance in T2DM is more closely related to compartment-specific lipid droplet remodelling, particularly expansion of subsarcolemmal lipid droplets, than to total IMCL alone (Nielsen et al. 2010, 2017; de Almeida et al. 2023). Accordingly, our Sudan Black B data should be interpreted as a comparative morphometric index of relative intracellular lipid storage, rather than a direct readout of lipotoxic lipid species or the subcellular lipid phenotype most closely linked to insulin resistance. These data reinforce the context dependence of IMCL, with muscle and fibre type shaping lipid storage patterns and adiposity contributing substantially to between-group differences (Goodpaster et al. 2001). The lack of clear group separation in the DIA and VL, together with the small, muscle-specific but fibre-type-independent changes, indicates that diabetes does not cause a uniform lipid overload across the musculature, but is superimposed on an existing gradient in which oxidative fibres store more lipid than glycolytic and hybrid fibres. The strong influence of BMI and the overall weak and regionally limited effects of HbA1c (with only a small isolated negative association in DIA type 1/2a fibres) support the view that adiposity and local oxidative propensity are the main drivers of IMCL in this cohort. Our BMI-adjusted models help account for group differences in adiposity, but they do not fully eliminate confounding by adiposity because the T2DM cohort had a substantially higher BMI and the study did not include a BMI-matched non-diabetic comparison group. This is particularly relevant for IMCL interpretation, as intramyocellular lipid accumulation is strongly influenced by adiposity and body fat status in human skeletal muscle. Therefore, the observed group differences may be interpreted as T2DM-associated alterations in the context of greater adiposity (Goodpaster et al. 2000; Moro et al. 2009). The slight negative association between age and IMCL in DIA and EXT may reflect reduced storage capacity or a shift towards greater lipid utilisation in chronically active respiratory tissues.
The 3D capillary analyses address limitations inherent to two-dimensional capillary indices. Two-dimensional measures derived from thin sections can be sensitive to fibre size, section orientation and regional sampling, which likely contributes to heterogeneous findings in human obesity and T2DM. Conventional stereological analyses have reported reduced capillarisation, unchanged indices or subtle changes that are difficult to interpret (Groen et al. 2014; Mortensen et al. 2019). In contrast, our 3D approach, combining confocal imaging, axial calibration, skeletonisation and vector-based reconstruction (Čebašek et al. 2010; Janáček et al. 2011), showed that microvascular architecture was largely preserved, with similar LVm, LL, LSf, MeanCap, branching density, and tortuosity between groups, and significant differences limited to DIA LVf and EXT anisotropy. A methodological limitation is that depth-uniform antibody penetration and fluorescence signal preservation were not quantitatively re-validated across the full 100 µm z-stack in the present dataset. However, our analysis was based on the established 3D confocal method of (Janáček et al. 2011), which identified penetration of fluorescent markers throughout thick human skeletal muscle sections as a required condition for reliable 3D capillary analysis and documented full-thickness penetration of the F8 endothelial antibody in originally 100-µm sections under optimised staining conditions. In the present study, all samples were processed at identical section thickness and with the same staining and imaging workflow, and linear Z-compensation was used to reduce depth-related attenuation. Nevertheless, residual depth-dependent signal loss or incomplete probe penetration could still lead to underestimation of absolute capillary length and branching measures and should therefore be considered when interpreting absolute 3D capillary metrics. Because the same protocol was applied across all muscles and both groups, such technical effects are less likely to account for the overall pattern of largely preserved capillary geometry, although subtle muscle-specific differences in labelling efficiency cannot be completely excluded.
Selective alterations in capillary supply metrics have been described in experimental obesity and diabetes, although the underlying driver varies by model and can include changes in fibre size, capillary remodelling or both (Poole et al. 2013; Gomes et al. 2017; Umek et al. 2021b). The DIA-specific LVf reduction in our study, therefore, likely reflects a modest imbalance between capillary length and fibre volume in a chronically loaded, oxidative muscle rather than frank vessel loss or network disorganisation. Covariate analyses further showed age-related simplification of the capillary network (lower tortuosity in SC and VL, shorter LL in VL), a positive association between BMI and LVf/LSf in the DIA, and DIA-specific associations of HbA1c within the T2DM group (shorter LL, higher MeanCap, trend to higher LS). Together, these patterns suggest that age and adiposity modulate quantitative capillary supply, particularly in the DIA, without disrupting overall microvascular topology, and that the DIA LVf deficit is a stable diabetes-related feature across the studied age range.
Across both groups, the intrinsic structural hierarchy among muscles was preserved: LVf followed the order DIA > SC ≈ EXT > VL, whereas fibre diameter exhibited the inverse pattern. This reciprocal relationship reflects a consistent design principle in skeletal muscle, whereby smaller oxidative fibres are supplied by proportionally denser capillary networks to optimise oxygen diffusion (Wüst et al. 2009) and parallels previous observations in healthy human muscle (Pollak et al. 2025) and in experimental obesity and diabetes models (Umek et al. 2021a). Its persistence suggests that mechanisms linking myofibre structure and capillary architecture remain largely intact in this cohort. Functionally, such structural stability may contribute to the relative preservation of respiratory and postural performance in individuals with T2DM, even though microvascular impairments and endothelial dysfunction are well documented in other tissues (Groen et al. 2014; Sörensen et al. 2016).
Given the chronic nature of diabetic tissue remodelling, the duration of diabetes is an important contextual exposure variable for interpreting the modest, muscle-specific phenotypes observed here. In our cohort, the recorded time since diagnosis was long (mean duration 16.5 ± 5.1 years), indicating long-standing diagnosed T2DM and providing context for interpreting the modest, muscle-specific phenotypes observed here. However, duration derived from clinical diagnosis is an imprecise surrogate for true glycaemic exposure, because T2DM commonly remains clinically unrecognised for years after onset of dysglycaemia (Harris et al. 1992). Accordingly, any inference that links ‘time since diagnosis’ to cumulative hyperglycaemic burden (or to the magnitude of structural differences) should be made cautiously. In addition, all individuals with T2DM were managed with oral antihyperglycaemic therapy (monotherapy or combination therapy), which may itself influence skeletal muscle metabolic and microvascular endpoints; for example, metformin has been shown in insulin-resistant humans to improve insulin-mediated skeletal muscle microvascular responsiveness alongside improved muscle glucose disposal, raising the possibility that treatment could have attenuated or modified diabetes-associated microvascular and lipid-related signals in this relatively well-controlled cohort (Jahn et al. 2022).
The main strengths of this study are its autopsy-based design, enabling simultaneous sampling of four anatomically and functionally distinct muscles from the same individuals, and the combined assessment of fibre type, IMCL and 3D capillary architecture. This design reduces interindividual variability, a major confounder in biopsy-based research, and provides a structurally coherent view of how T2DM affects the muscular system. However, several limitations should be acknowledged. The cohort consisted solely of older male individuals, limiting generalisability to women and younger age groups, in whom sex hormones and physical activity patterns may influence muscle phenotype and vascularisation. The post-mortem, cross-sectional design precludes assessment of dynamic perfusion, mitochondrial function or muscle performance, and structural preservation therefore cannot be directly equated with preserved functional capacity (Eržen et al. 2018). Mitochondrial enzyme histochemistry was not performed because, in post-mortem muscle, interpretation of enzyme-based histochemical readouts would require dedicated validation and standardisation of post-mortem interval-related and tissue-handling effects across all four muscles, which was beyond the scope of the present study; accordingly, the present data do not directly address mitochondrial alterations reported in skeletal muscle in T2DM (Kelley et al. 2002). An additional limitation of the autopsy-based design is that agonal physiology cannot be fully controlled. Pre-mortem hypoxia and hemodynamic instability can affect endothelial barrier function and capillary permeability, and hypoxia can also alter skeletal muscle fatty acid handling and intracellular lipid composition (Morash et al. 2013). Post-mortem tissue studies also indicate that death-related and post-mortem factors may influence biological readouts in a tissue-specific manner (Ferreira et al. 2018). Although all samples were collected within 24 h post-mortem, and tissues were grossly well preserved, subtle autolytic or fixation-related effects cannot be fully excluded.
Furthermore, while cause-of-death data were available for all cases, agonal duration and peri-mortem treatment and medication data were not available in a standardised form, and cause-of-death profiles were not matched between groups. Therefore, the IMCL and microvascular findings should be interpreted as muscle-specific alterations observed in well-characterised autopsy cohorts, in which diabetes-related remodelling may be accompanied or modulated by agonal and peri-mortem factors. The T2DM group was also not BMI-matched to controls, so despite statistical adjustment for BMI, residual confounding by adiposity cannot be excluded, especially for IMCL-related outcomes and potentially for microvascular phenotypes linked to obesity-associated metabolic dysfunction. Finally, molecular markers of angiogenesis, oxidative metabolism and inflammation (e.g. VEGF, PGC-1α, TNFα) were not measured, so mechanistic interpretations remain hypothesis-generating.





Comments (0)