
Remember me
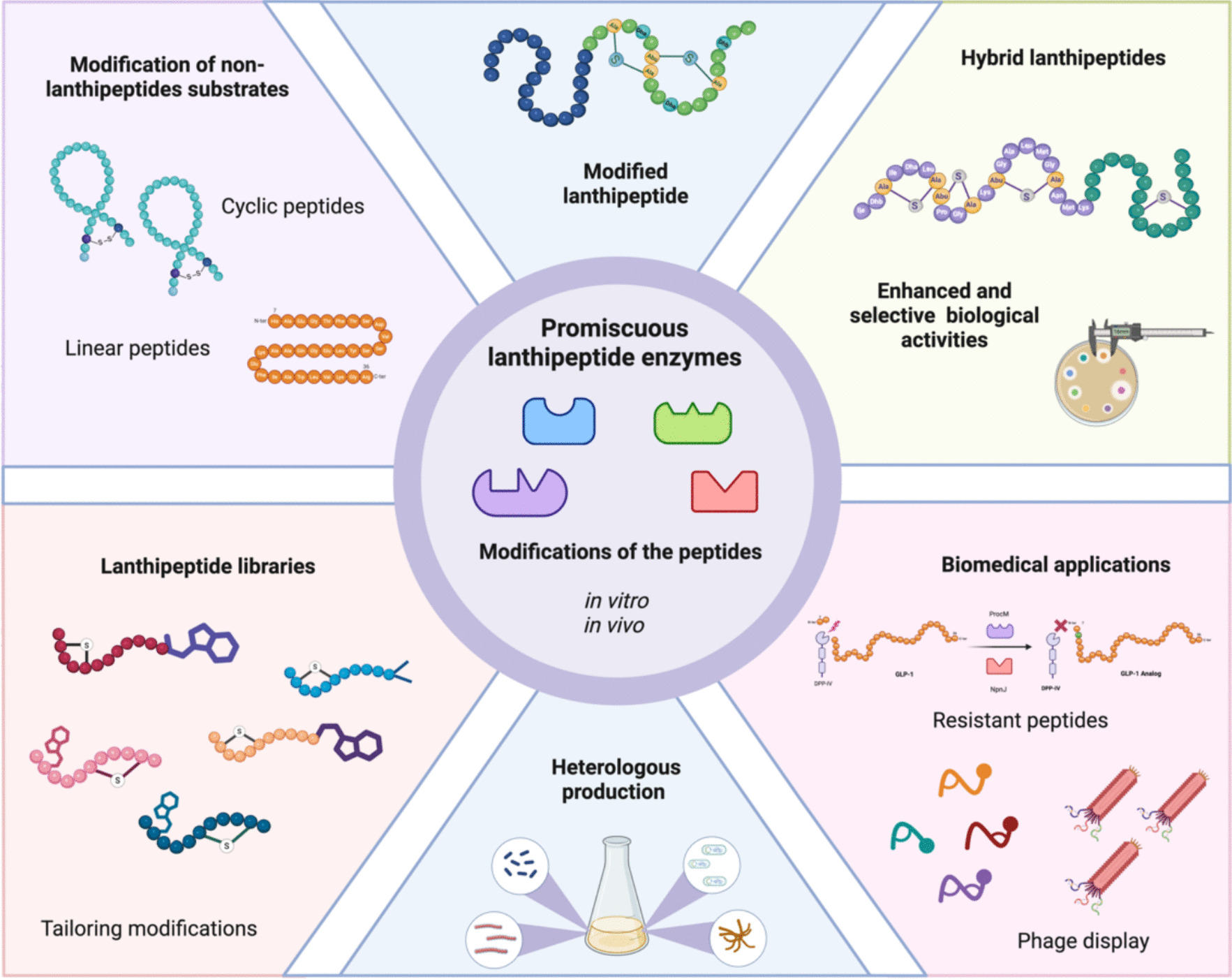

Gram-positive bacteria mainly produce lanthipeptides, but bioinformatic analyses reveal many biosynthetic gene clusters for lanthipeptides in Gram-negative bacteria and archaea. This is notable given the limited number of characterized lanthipeptides in these groups (van der Donk and Nair 2014; Barbosa et al. 2015; Caetano et al. 2020; Walker et al. 2020; Castro et al. 2021; Ramírez-Rendón et al. 2023; Roblero-Mejía et al. 2024). These peptides are noted for their biological properties, particularly their antimicrobial activity against multidrug-resistant bacteria (MDR), such as Methicillin-resistant Staphylococcus aureus, vancomycin-resistant enterococci (VRE), and various clostridia strains (Singh et al. 2020; van Staden et al. 2021; Shi et al. 2022; Deisinger et al. 2023; Wang et al. 2025). In addition to their well-documented antimicrobial effects, lanthipeptides have also been found to possess antiviral (Blockus et al. 2020; Fu et al. 2021; Singh et al. 2023), antifungal (Mohr et al. 2015; Singh et al. 2023), and other significant biological activities (Iorio et al. 2014; Han et al. 2022; Wang et al. 2023). Several studies have explored even their potential as diagnostic molecules due to their ability to bind to cell membrane components, such as phosphatidyl ethanolamine in eukaryotic cells (Luo et al. 2016). Nisin (NIS) is a lanthipeptide that has been widely used as a food preservative for over fifty years due to its strong activity against food pathogens, including Listeria monocytogenes (Gharsallaoui et al. 2016; Ibarra-Sánchez et al. 2020). This knowledge positions lanthipeptides as promising candidates for various clinical and biotechnological applications (Ramírez-Rendón et al. 2023). These peptides are categorized into the ribosomally synthesized and post-translationally modified peptides (RiPPs) and are distinguished by the presence of lanthionine (Lan) and methyl-lanthionine (MeLan) rings in its structures (Repka et al. 2017; Hudson and Mitchell 2018). The precursor peptide (PP) is divided into two regions: the leader peptide and the core region, and it is encoded by a biosynthetic gene cluster (BGC) (van der Donk and Nair 2014; Repka et al. 2017). The biosynthesis of lanthipeptides starts with the production of the peptide by ribosomes. This is followed by the binding of the leader peptide (LP) to specific biosynthetic enzymes. These enzymes then carry out post-translational modifications on the core peptide (CP). Finally, a protease cleaves the LP, resulting in a mature peptide that is responsible for the compound's biological activity (Fig. 1a). Currently, six distinct classes of lanthipeptides have been identified, each characterized by specific enzymes that facilitate dehydration and cyclization reactions to form Lan and MeLan rings (Xu et al. 2020; Lagedroste et al. 2020; He et al. 2023) (Fig. 1b). These enzymes display broad substrate specificity, often referred to as substrate tolerance or substrate promiscuity. This concept is a form of enzymatic promiscuity, where a single enzyme can catalyze the same chemical reaction on different substrates (Hult and Berglund 2007). The term was first used in the context of lanthipeptide biosynthesis over two decades ago, specifically in studies of the NIS biosynthetic cluster, which was employed to install rings in non-lanthipeptide substrates (Kluskens et al. 2005). This concept was later supported by other biosynthetic enzymes, such as lacticin 481 synthetase (Kuipers et al. 2004; Chatterjee et al. 2006; Zhang and van der Donk 2007; Rink et al. 2007). Since the discovery of prochlorosins (Li et al. 2010), a class II lanthipeptide composed of 29 precursor peptides (PPs), new evidence has emerged showing that biosynthetic enzymes can modify multiple PPs with varying structural features. These enzymes are referred to as promiscuous enzymes (Tang and van der Donk 2012), highlighting their ability to work with a relaxed range of substrates. Both terms are currently used to describe the ability of these enzymes to catalyze the same reaction (such as dehydration or cyclization) across different peptides, even if those peptides do not belong to the original cluster. This characteristic is often associated with the LP involved in the process of enzymatic recognition (Majchrzykiewicz et al. 2010; van Heel et al. 2016). Additionally, the presence of motifs and secondary structures is crucial for the correct processing of the products, although this role varies according to the lanthipeptide class (Plat et al. 2013; Hegemann and van der Donk 2018; Wiebach et al. 2020). A conformational model has been proposed, where the LP binds to the biosynthetic enzyme, thereby improving the affinity to CP due to the observed low affinity of these enzymes to CP in various systems. This feature allows the modification of different substrates (Levengood et al. 2007; Oman et al. 2012; Abts et al. 2013; Thibodeaux et al. 2015; Rahman et al. 2020). Therefore, understanding these characteristics is essential for various applications. From the modification of mutational cognate-peptides (Zhang and van der Donk 2007) to forming rings on medically important peptides such as angiotensin and erythropoietin (Kluskens et al. 2005) to improve their stability, the use of promiscuous enzymes allows the fast production of new and improved peptides. A study conducted in 2016 used NIS machinery to produce five new lanthipeptides with high antibacterial activity without the need to be modified by their cognate enzymes, reducing challenges by using already known systems (van Heel et al. 2016). Even, the broad substrate tolerance of ProcM has been used to produce lanthipeptide libraries. The search for inhibitors of HIV p6 protein in human cells, led to the creation of a lanthipeptide library of 106 peptides using the same LP (Yang et al. 2018), supporting the great usefulness of promiscuous enzymes to produce new peptides of biotechnological importance. At the same time, we cannot ignore the usefulness of tailoring enzymes to diversify to lanthipeptides. The protease ElxP and the short-chain dehydrogenase ElxO, from epilacin 15X biosynthesis, demonstrated relaxed substrate specificity to cut NIS and reduce lactocin S (a class I lanthipeptide), respectively, supporting the fact that tailoring enzymes can also be promiscuous enzymes (Ortega et al. 2014).
Fig. 1
The processing of lanthipeptides: a The biosynthetic pathway of lanthipeptides; b the general mechanisms of lanthionine ring formation across all known lanthipeptide classes today
In this review, we summarize the latest research advancements in the use of promiscuous enzymes to produce new natural products and their potential biotechnological applications, with a specific focus on lanthipeptides. We also highlight key properties of the biosynthetic enzymes involved in lanthipeptide production.
LanB and LanC enzymesBiosynthesis of class I lanthipeptides is catalyzed by two primary enzymes: a dehydratase, LanB, and a cyclase, LanC. In general, these enzymes are functional in several heterologous hosts like Escherichia coli, several actinobacteria, Lactococcus sp., Bacillus, and in vitro systems (Foulston and Bibb 2010; Sherwood et al. 2013; Liu et al. 2020; Zhao and Kuipers 2021; Chen et al. 2021; Chen and Kuipers 2022; Kaweewan et al. 2022; Lee et al. 2023). Most studies on the enzymatic mechanisms of this class of lanthipeptides have concentrated on the NIS enzymes, NisB and NisC. This system has advantages over other enzyme systems, such as those found in actinobacteria (for instance, the microbisporicin biosynthetic system), because it effectively catalyzes dehydration and cyclization reactions without the need for NisB or NisC to function in concert (Ortega et al. 2016; Repka et al. 2018). Importantly, both NisB and NisC require a LP for catalyzing ring formation. Research on NisB and NisC has highlighted the significance of the FxLx motif in the LP, as well as the STKD (−12 to −19) and PR (−2 to −1) motifs for binding to the precursor peptide (Plat et al. 2011; Mavaro et al. 2011; Khusainov et al. 2013; Repka et al. 2018). Interestingly, functionality with leaderless peptides has been demonstrated in vivo when an antimicrobial product was produced in Lactococcus lactis NZ9000. This reaction was enhanced when the LP was expressed in trans, resulting in fewer premature nisin molecules and increased antibacterial activity (Khusainov and Kuipers 2012). These findings highlight the crucial role of the LP in binding biosynthetic enzymes, particularly NisB, which exhibits a greater affinity for the LP compared to NisC. Additionally, in this system, specific mutations in the LP are more easily tolerated (Rink et al. 2005; Plat et al. 2011; Abts et al. 2013). This flexibility allows for the incorporation of identification and purification tags on the N-terminal side, even for larger peptides.
Recently, it was reported that fluorescence proteins such as mCherry red fluorescent protein are well tolerated by the nisin and clausin machinery, as well as the green fluorescent protein (GFP), which was added to LP of both NIS and Pep5 in E. coli, where the peptides were successfully processed and the mature peptides remained active (van Staden et al. 2019; Van Zyl et al. 2023).
NIS was the first lanthipeptide identified in lactic acid bacteria (LAB) (Delves-Broughton et al. 1996; Field et al. 2023). It is well-researched and documented. Due to its antibacterial properties against Gram-positive strains, it serves as a prototype for lanthipeptide engineering. Regarding its biosynthetic pathway, although the NisB and NisC enzymes are mainly involved in the production of NIS, other proteins, such as the ATP-binding cassette (ABC) transporter (NisT) and the subtilisin-like serine protease (NisP), are also related to the export and proteolysis of mature NIS (Cheigh and Pyun 2005; Lubelski et al. 2008). On the other hand, proteins related to resistance and immunity are also involved in the production of this lanthipeptide. The NisR and NisK are the key regulators in triggering NIS synthesis (Engelke et al. 1994; Qiao et al. 1996). In this sense, the NIS-controlled expression system (NICE) is utilized for several antimicrobial peptides, which use the NIS biosynthetic enzymes to induce lanthipeptides. The first characterized strain for NIS production was Lactococcus lactis subs. cremoris MG3163 (later re-named as NZ900). It was described that the insertion of nisR and nisK genes into the genome of the strain triggered the production of a cloned product in a plasmid with NisR promoter, using lower amounts of NIS (0.1–5 ng/mL) (Mierau et al. 2005; Mierau and Kleerebezem 2005). Likewise, other similar systems, controlled by subtilisin, are not limited to NIS utilization. Thus, it can be used by Bacillus subtilis (SURE) for the production of β-glucuronidase and green fluorescent protein (GFP) reporters, utilizing the promoter PspaS, and the regulatory genes spaR and spaK (Bongers et al. 2005). The NICE system was utilized to produce a wide variety of biomedical and industrial products, as well as specific class I lantibiotics (antibiotics containing lanthionine rings) to produce bioactive peptides (Mierau and Kleerebezem 2005; Montalbán-López et al. 2017; Sandiford 2020; Frelet-Barrand 2022). Over the years, these proteins have demonstrated great flexibility in accepting non-wild substrates, provided that the PL sequence in the peptides is preserved, with variations allowed at specific sites within the CP (Table 1). For instance, the antimicrobial peptides (AMPs) thanatin and rip-thanatin, effective against Gram-negative bacteria, are sulfur bond-containing compounds produced by the insects Podisus maculiventris and Riptortus pedestris, respectively. These peptides were used as peptide backbones to create a peptide incorporating a MeLan ring using the NICE system in L. lactis NZ900, forming thanacin and ripcin (Zhao and Kuipers 2021). Another example is the hybrid peptide known as RipC, which is created by combining the lipid II-binding region of NIS (comprising the first 20 amino acids) with ripcin (Fig. 2a). This peptide has demonstrated effectiveness against MRSA and Acinetobacter baumannii, displaying selective binding to lipid II (Zhao and Kuipers 2021) (see Fig. 2b).
Table 1 Recent reports on peptides produced by class I and II enzyme promiscuity. In general, their bioactivity and stability are improvedFig. 2
Comments (0)