
Remember me

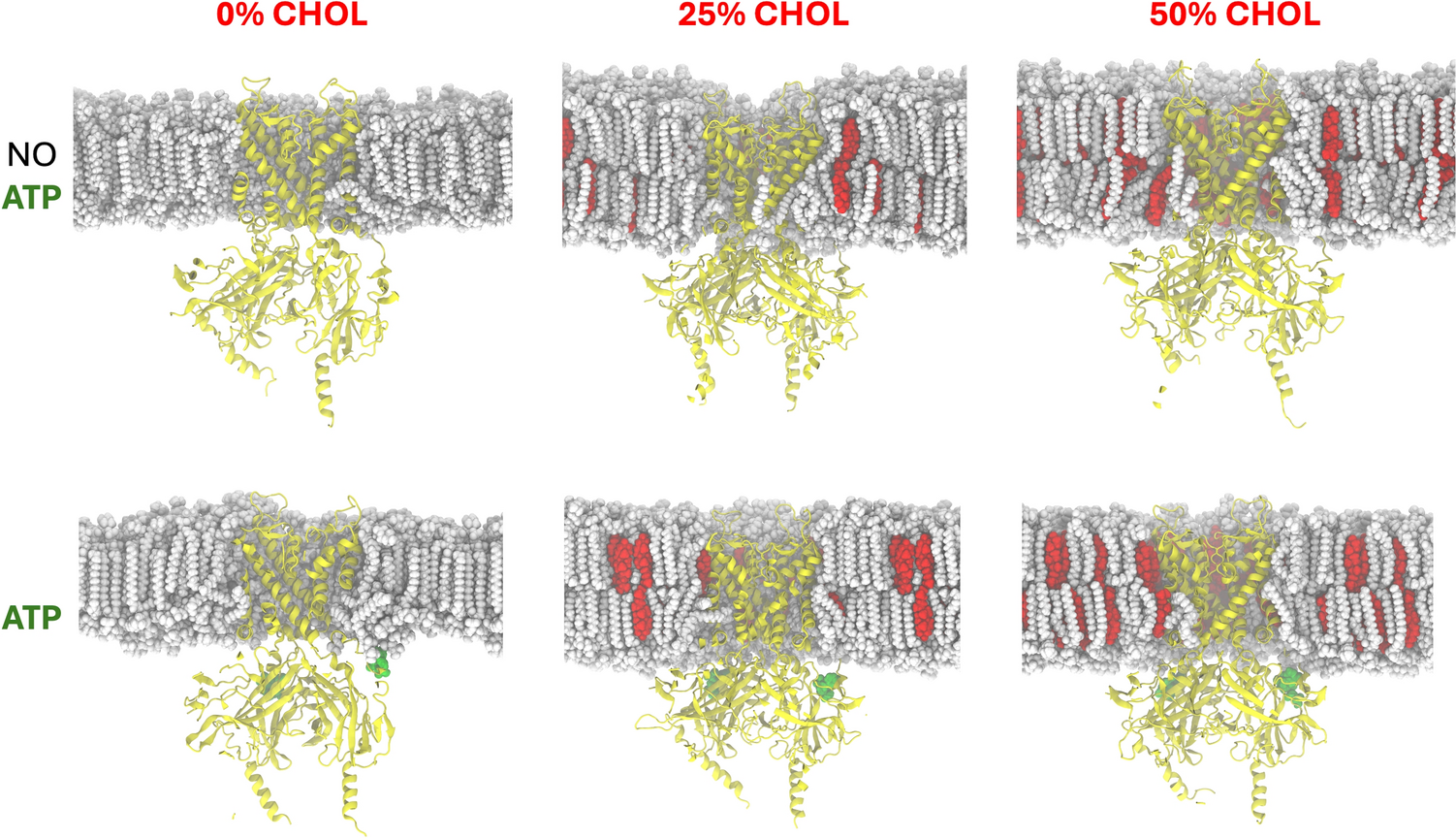
This study aimed to determine whether and to what extent the channel-forming agent nystatin passes through the ergosterol-containing phospholipid membrane, which serves as a model for the cell membrane. For this purpose, both monovesicular giant unilamellar POPC vesicles (GUVs) and giant multivesicular POPC vesicles (MVVs), which consist of outer (outGUVs) and inner (inGUVs) vesicles, were exposed to nystatin and their rupture times were compared. By comparing the rupture times of GUVs and inGUVs of the same size, the passage of nystatin through the vesicle membrane could be deduced. MVVs can also be used to study the ability of other molecules to passively cross the lipid membrane. In the case of amphipathic polypeptides, their passage was determined by the fluorescence signal that occurs due to the passage of the corresponding dye (Wheaten et al. 2013).
Explanation of Vesicle RupturesA brief explanation of vesicle rupture (e.g., rupture of GUV, outGUV, or inGUV) induced by a channel-forming agent should be given. When the vesicle membrane is exposed to sufficiently high concentrations of the channel-forming agent nystatin, size-selective transmembrane channels are formed (Holz and Finkelstein 1970; Katsu et al. 2008). A selective passage, i.e., a higher flux of smaller glucose molecules compared to larger sucrose molecules, is induced (Kristanc et al. 2014). Since in our experiments the glucose molecules are outside the vesicle and the sucrose molecules are in the inner solution, the number of sugar molecules and, consequently, the sugar concentration inside the vesicle increase. As a result, a net flux of water into the vesicle is induced and the volume of the vesicle increases. When the area of the vesicle reaches its critical value, the vesicle ruptures (Koslov and Markin 1984; Idiart and Levin 2004).
In addition to increasing the permeability for glucose, nystatin also significantly increases the permeability of the membrane for water. Both increases are necessary for the large influx of water (Kristanc et al. 2014). This is because a considerable influx of water is required for vesicle rupture, in which the membrane is completely disintegrated.
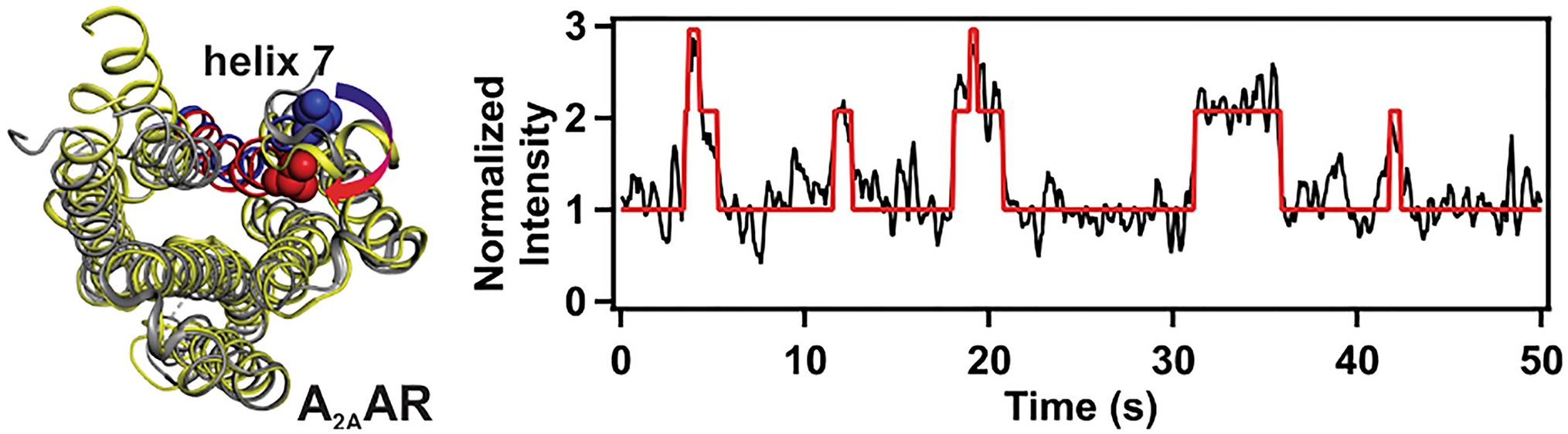
Experimental Evidence of Nystatin PassageComparable rupture times of GUVs and outGUVs of the same size show, according to the previous discussion, that the nystatin channels are formed in both types of vesicles over the same time course (Fig. 2). In contrast, the experimental data show that the rupture times of GUVs and inGUVs with the same size are significantly different, indicating a different time course of the nystatin channel formation in their membranes. In addition, their times in the bulk solution, i.e., in the 250 or 500 \(\mu\)M nystatin concentration, are significantly different. As can be seen in Fig. 4, the time between the exposure of the GUV to the nystatin solution and its rupture is significantly longer than the time between the rupture of the outGUV and the rupture of the inGUV, i.e., the time during which the inGUV is exposed to the bulk solution. The earlier rupture of the inGUVs indicates that, unlike the GUV, the inGUV has already incorporated nystatin into the membrane before it is exposed to the bulk solution, which is the reason why the inGUV ruptures earlier than the GUV. However, this incorporation of nystatin into the membrane of the inGUV is only possible if nystatin molecules pass through the membrane of the outGUV into its interior and bind to the membrane of the inGUV before the outGUV ruptures.
Also, in any of the experiments the rupture of the outGUV did not occur simultaneously or later than the rupture of the corresponding inGUV, even if the radius of the inGUV is much smaller than the radius of the outGUV of the same MVV. This indicates that the nystatin molecules cannot pass the membrane of the outGUV unhindered. In summary, the assumptions that nystatin does not pass through the membrane or that it passes through the membrane instantly, are therefore deficient.
The nystatin passage is also supported by the comparison of the radius increase between inGUVs and GUVs with radii smaller than 14 \(\mu\)m (Fig. 6). For all tested nystatin concentrations and ergosterol contents, the inGUVs initially show a larger increase in radius immediately after the exposure to the bulk solution than the corresponding GUVs (Fig. 6). The increase in vesicle radius can only be larger if the portion of nystatin molecules in the membrane, causing the greater influx of glucose and water, is greater. The glucose and water molecules can therefore pass through the membrane of the inGUV to a greater extent after rupture of the corresponding outGUV, which indicates that the nystatin is already incorporated into the membrane of the inGUV when the outGUV ruptures.
It should be noted that the average radius shown in Fig. 6 reflects the volume of the vesicle, as the vesicle volume is, to a first approximation, proportional to the cube of its radius. However, the vesicle radius is less relevant for determining the membrane area because an increase in membrane area at constant volume primarily decreases lateral tension and, accordingly, increases membrane fluctuations, which are not directly reflected in an increase in vesicle radius (Seifert 1997).
Additionally, before the outGUV ruptures, there is an influx of glucose through its channels into its interior, which leads to a slightly higher sugar concentration in the solution surrounding the inGUV. This tends to reduce the volume of the inGUV if no nystatin is bound to its membrane. In this case, due to the lower volume of the inGUV at the exposure to the bulk solution, it would take longer for the inGUV to rupture than a GUV of the same size. In contrast, we observe in the experiments that the mean time of the inGUV in the bulk solution is shorter than the mean time of the GUV of the same size in the bulk solution. This conclusion is in favor of the passage of nystatin through the membrane.
The results also show that the radius increases of vesicles exposed to the nystatin solution are considerably large (subsection Increase of the vesicle radius 3.3). The increases could, to some extent, be explained by the excess membrane area, i.e., the reduction of membrane undulations, since our vesicles are not initially tense. Our control measurements showed an approximate 8% increase in membrane area due to reduced membrane undulations and membrane stretching, which corresponds to the contribution of thermal fluctuations and membrane elasticity in lipid membranes (Seifert 1997; Evans et al. 2013). In addition, other effects contributing to increases in membrane area cannot be neglected, including: (i) binding and pore formation by nystatin, (ii) methanol binding, and (iii) changes in the membrane’s stretching constant and lysis tension due to nystatin and methanol binding. Our control measurements showed an approximate 3.3% increase in membrane area due to the effect of methanol, leading to a total increase of about 12% in membrane area without accounting for the effects of nystatin. This leaves approximately 24% of the area increase ascribed to nystatin binding and pore formation, as well as to the changes in the membrane’s stretching constant and lysis tension caused by the intercalation of nystatin and methanol. The density of nystatin pores must be sufficiently high to cause vesicle rupture—approximately one pore per 100 lipid molecules (Kristanc et al. 2014)—which could result in an additional 7% increase in vesicle area. We may also speculate that membrane thinning, a phenomenon similar to that induced by high methanol concentrations, could occur around the nystatin pores, thereby increasing surface area (Simon and McIntosh 1984; Zeng et al. 1993; Kristanc et al. 2012). Nevertheless, it must be emphasized that our experimental determinations of the radius increase, with a standard deviation of approximately 40%, are not accurate and, therefore, are not used as the primary evidence for the passage of nystatin molecules through the membrane. However, these results highlight the substantial impact of nystatin on membrane behavior.
Proposed Mechanism of Nystatin PassageAccording to the well-accepted model of nystatin channel formation, nystatin aggregates interact with the phospholipid membrane and dissolve, allowing monomers or small oligomers to bind horizontally to the outer layer of the phospholipid bilayer (Zemel et al. 2005; Cohen 2010; Szomek et al. 2021). At a sufficient membrane density, these monomers or small oligomers undergo oligomerization and reorientation perpendicular to the membrane surface (Recamier et al. 2010), which is facilitated by the amphipathic nature of nystatin. Once embedded, they continue to oligomerize, forming transmembrane channels (Kleinberg and Finkelstein 1984; Zemel et al. 2005). The number of these channels and the number of nystatin monomers composing the individual channel can vary over time.
Fig. 7
Schematic representation of the mechanism that enables the passage of nystatin molecules through the POPC membrane. A passive net flux of nystatin molecules across the membrane from the side with a higher nystatin concentration to the side with a lower concentration is denoted by the arrows. This flux is characterized by the binding of aggregates, the dissociation of monomers or small oligomers from aggregates, the formation of channels, the dissociation of single molecules from the channels, the formation of oligomers and aggregates, and the unbinding of molecular aggregates. The representation corresponds to two possible ways in which nystatin spans the lipid bilayer: by a tandem insertion into both monolayers (a) or by a conformational thinning of the membrane (b)
Based on this model, the following mechanism is proposed for the passage of nystatin through the phospholipid bilayer: some monomers can dissociate from the individual channels and can move into the solution on both sides of the membrane (Fig. 7). Consequently, a passive net flux of nystatin molecules is induced across the membrane from the side with a higher nystatin concentration to the side with a lower concentration. The proposed mechanism predicts that the passage of nystatin molecules through the membrane only occurs when the nystatin channels are formed in the membrane.
The proposed mechanism is supported by the experiments performed at high nystatin concentrations (above 250 \(\mu\)M), where the formation of nystatin channels leads to the ruptures of the vesicles. However, previous studies have shown that nystatin channels are also formed at much lower nystatin concentrations, below 25 \(\mu\)M (Recamier et al. 2010). We postulate that the mechanism for the passage of nystatin molecules through the membrane may be operational at lower nystatin concentrations, where channel formation still occurs, although vesicle ruptures are absent or occur far less frequently. These lower nystatin concentrations are often physiologically relevant. Nystatin concentrations in internal organs such as the kidneys and liver typically remain much lower—in the range of 1–10 \(\mu\)M (Semis et al. 2012). In contrast, after intravenous application, nystatin concentrations can reach up to 35 \(\mu\)M in the bloodstream, and after topical application, levels may reach up to 200 \(\mu\)M in the upper layers of the skin or mucosal surfaces (Groll et al. 2003; Semis et al. 2012). It is to be noted that the passage of nystatin molecules by this mechanism is impossible at nystatin concentrations where no transmembrane channels are formed due to an insufficient areal density of nystatin molecules on the membrane surface (Recamier et al. 2010). Therefore, our conclusions do not contradict the literature where no nystatin permeability in lipid membranes was observed.
Role of Ergosterol Content and Nystatin ConcentrationThe role of ergosterol lies in enhancing the solubility of nystatin in the membrane (Szomek et al. 2021). In particular, membrane structure, which varies with sterol content, is involved in the solubility of nystatin aggregates in the membrane (Recamier et al. 2010), but the nystatin channels do not require sterol for their formation. This effect could be due to complex, non-linear dynamics, as suggested in previous studies (González-Damián and Ortega-Blake 2010). Therefore, the role of sterol-dependent membrane structure is crucial for polyene activity. However, our experimental results showed nonsignificant differences in average rupture times and average radius increase between vesicles with 15 and 45 mol% ergosterol membrane contents at the same nystatin concentration, with variations falling within the standard deviation of the mean. It must be taken into account that the scatter of the measured rupture times is large. In addition to a data scatter due to biological variability, the scatter due to the random occurrence of ruptures appears (Chabanon et al. 2017).
Further experimental evidence supporting the passage of nystatin through the phospholipid membrane is the significantly smaller differences in the times in the bulk solution between 250 and 500 \(\mu\)M concentrations for the inGUVs compared to the GUVs (Figs. 4 and 5). Namely, because nystatin passes through the phospholipid membrane of the outGUVs, the key distinction between GUVs and inGUVs is that inGUVs already have nystatin incorporated into their membranes at the beginning of their exposure to the bulk solution, whereas this is not the case for GUVs. The relatively small difference in the times in the bulk solution between 250 and 500 \(\mu\)M concentration for the inGUVs thus suggests that the process of nystatin incorporation into the membrane of the inGUVs is largely complete at both 250 and 500 \(\mu\)M nystatin concentration before the outGUV ruptures. It could be speculated that after the rupture of the outGUV, i.e., after the exposure of the inGUV to the bulk solution, the rupture times of inGUVs are mainly influenced by glucose and water influxes through the already formed nystatin channels. In contrast, the GUVs that start without bound nystatin when exposed to the bulk solution show a stronger effect of nystatin concentration on channel formation, resulting in significantly shorter rupture times at 500 \(\mu\)M than at 250 \(\mu\)M concentration.
Accordingly, a lower limit for the membrane permeability of nystatin can be estimated on the basis of the rupture times (Fig. 3), assuming that glucose can pass through the membrane unhindered (Supplementary Information). This limit is in the order of magnitude 0.1 \(\mu\)m/s, considering that the permeability of the membrane for nystatin (P) is defined by the equation, \(j = P\Delta c\), where j is the particle flux density through a membrane and \(\Delta c\) is the concentration difference between the two sides of the membrane (Fick’s law).
Comments (0)