
Remember me
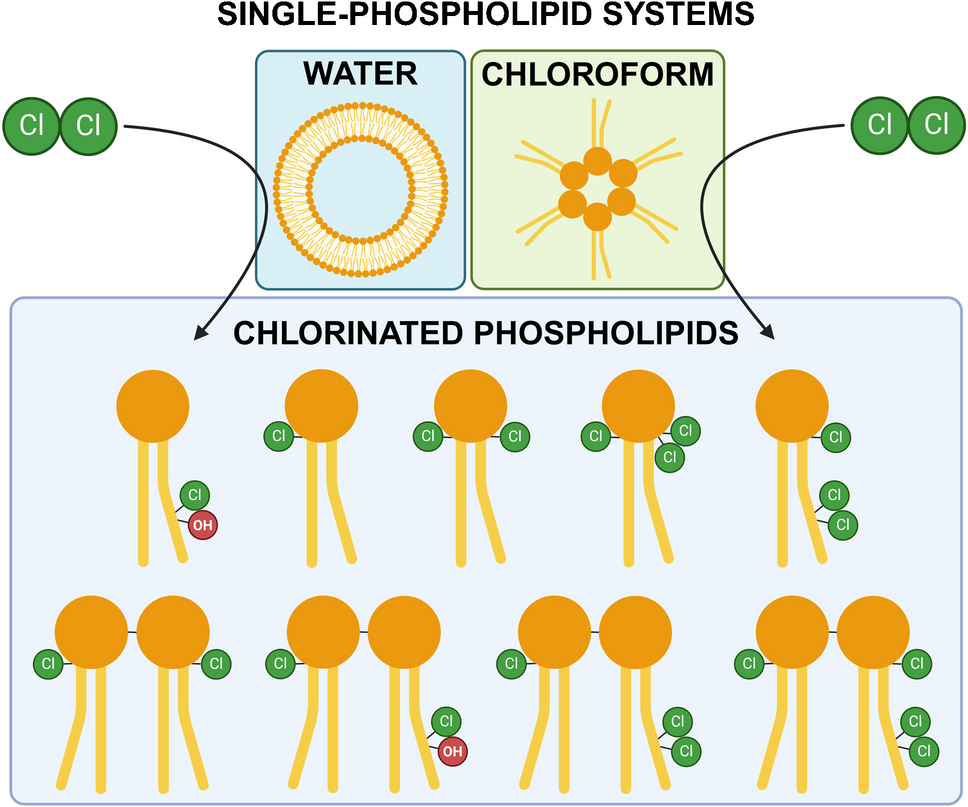

The POPC samples chlorinated as vesicles in water were first analyzed with LC–MS and LC-HRMS using a C18 column in positive polarity, before further analysis with HILIC separation. Chlorinated PCs were not detected in the untreated control sample. Five chlorinated PCs were found in the chlorine-exposed sample: PC chlorohydrin (PC HOCl), PC dichloride (PC D), and analytes with theoretical accurate m/z values of 794.54611 (PC A), 777.56560 (PC B), and 803.55139 (PC C) (Table 1). Evaluation of the isotopic patterns of PC A–C revealed that PC A and B contain one chlorine atom and PC C two chlorine atoms (Fig. S10–S12). The 13C peaks of PC B and C differed by approximately 0.5 mass units from the monoisotopic ion peak (Fig. 2, and Fig. S11 and Fig. S12 in the Electronic Supplementary Material (SM)), indicating that the analytes are doubly charged.
Table 1 Abbreviations of the chlorinated lipids, their predicted formula, and theoretical and measured m/z. Masses corresponding to the lipids marked with an asterisk (*) were detected in the chlorinated lung sampleFig. 2
TIC of a POPC sample chlorinated in water separated by HILIC and the isotopic patterns of PC A (left) and PC C (right). The chlorinated PCs elute in three structure-dependent groups: PCs chlorinated in the glycerol backbone (e.g., PC A), PCs chlorinated in the acyl chains (e.g., PC D), and dimeric chlorinated PCs (e.g., PC C). The isotope peaks in the isotopic pattern of PC C are 0.5 units apart, which is typical for doubly charged ions
A chlorinated PC with an accurate mass of 794.54611 has previously been reported in POPC samples incubated with HOCl [17]. Based on the fragmentation pattern, the authors suggested that the analyte was a POPC chlorinated in one of the acyl chains without opening the double bond. Here, LC–MS/HRMS with stepped HCD fragmentation was used to fragment PC A–C to obtain structural information. All three lipids produced typical phosphocholine fragments (m/z 86.09643, 98.98417, 104.10699, 124.99982, and 184.07332, proposed structures in Fig. 3) [25], indicating that the chlorine substituent is not located in the headgroup. This is also supported by the chlorinated fragments consisting of the glycerol backbone and one or two acyl chains (e.g., m/z 977.61184, 794.54611, 611.48006, 556.31644, and 373.25040) (Tables S3, S5, and S6; Fig. S21–S23 in the SM). Interestingly, PC A and PC C produced a fragment ion with an m/z of 202.03943 (Fig. 3), corresponding to the phosphocholine headgroup in which one of the hydroxyls in the phosphate has been replaced with a chlorine atom (Tables S3 and S6; Fig. S31 and S33 in the SM). This fragment is presumably produced by re-arrangement reactions during the collisional fragmentation. For the re-arrangement reaction to occur, the chlorine atom must be in close proximity to the phosphate, i.e., either in the headgroup or in the glycerol backbone. Since PC A and C produce the aforementioned phosphocholine fragments, it is unlikely that the chlorine substituent is in the headgroup. PC A also produced fragment ions with m/z 239.23694, 263.23696, and 199.03678. The first two mentioned are ions produced in the fragmentation of the un-chlorinated acyl chains. The last ion mentioned could potentially have the molecular formula C6H12ClO5+, which corresponds the chlorinated glycerol backbone (Fig. 3).
Fig. 3
Proposed structures of selected fragments of the chlorinated PCs
Based on the aforementioned and other fragments of PC A–C (and the other detected chlorinated PCs discussed in the “POPC chlorinated in chloroform” section), we propose that the chlorine substituent is located in the glycerol backbone, which differs from previously made conclusions on the structure of the chlorinated PC with the m/z value of 794.54611 [17]. This hypothesis is supported by previous studies where chlorination of ethers resulted in the addition of chlorine in the α- and β-positions. While the α-carbon of the carbonyl can also be chlorinated [26], the glycerol backbone might be a more available reaction site, hence leading to the observed products. Selected fragments of the chlorinated PCs are presented in Fig. 3. The exact position of the chlorine atom(s) in the glycerol backbone cannot be concluded from the data available. However, multiple peaks were detected for many of the analytes, suggesting that multiple structural isomers of the chlorinated PCs were formed during the chlorination.
Based on the two charges and the fragmentation (e.g., m/z 489.30972 = PC A with an additional phosphocholine headgroup), we propose that PC B and C are chlorinated, dimeric peroxy-diphospholipids in which one of the lipid monomers is PC A. The second lipid in the dimers is POPC or PC HOCl in PC B and PC C, respectively.
The structures of PC HOCl and PC A–D (and PC E–L, which are discussed in the “POPC chlorinated in chloroform” section) are presented in Fig. 4. The isotopic patterns of the analytes are presented in Fig. S9–S13, and the fragmentation patterns of the analytes can be found in Tables S1, S3, S5, S6, and S8 in the SM. The proposed structures for the detected fragments are presented in Fig. S30–S34 in the SM.
Fig. 4
Proposed structures of PC HOCl and PC A–L, as detected by LC-HRMS. The chlorine substituent(s) may be located in any of the carbons in the glycerol backbone
To obtain further structural information, the samples were also analyzed with HILIC LC-HRMS. In C18, retention is mainly based on hydrophobicity; hydrophobic analytes are retained more strongly than hydrophilic analytes. The hydrophobicity of PCs is determined by the lengths and degrees of unsaturation of the alkyl chains [27]. The retention of analytes in HILIC columns is more complicated, but generally polar and ionic compounds are retained more effectively [28]. The polarity of PCs is mainly determined by the phosphocholine headgroup [27]. PC B and C show the strongest retention on HILIC, presumably because they both have two charged cholines which like to interact with the hydrated zone and charges of the stationary phase. The retention times suggests that PC A is the least polar/ionic of the four chlorinated PCs. An electronegative chlorine atom in the glycerol backbone might decrease the net-polarity of PC A, e.g., by attracting the quaternary amine or by changing the charge distribution in the headgroup. This can, in turn, result in changes in the PCs’ interactions with solvent molecules and the stationary phase and shifting of the retention times. Further research is needed to determine how the chlorine modification in the glycerol backbone affects the structure and properties of PCs. Nevertheless, the charge and chlorine substituent position–dependent retention in HILIC separation were useful in determining the structure of the chlorinated PCs and PEs discussed in the “POPC chlorinated in chloroform” and “Alkaline and enzymatic degradation of chlorinated POPC” sections.
Experiments using ESI negative polarity scanning were also performed with C18 LC–MS. Of the chlorinated analytes, only PC HOCl and PC D were detected as negative ions ([M-CH3]−, m/z 796.5 and m/z 814.5, respectively, data not shown). In the case of the glycerol-chlorinated lipids, the inability to produce negatively charged ions may have been due to changes in ionizability, caused by the very electronegative chlorine atom near the headgroup. Similarly to un-chlorinated PCs, PC HOCl and PC D (which contain chlorine atoms only in their acyl chains, see Fig. 4) were negatively ionized, through a loss of a methyl group and subsequent neutralization of the quaternary ammonium choline [25].
For further structural investigation of the position of the chlorine substituents and the dimeric lipids, a sample was prepared for 1H nuclear magnetic resonance (NMR) analysis. A 1 mL sample containing 10 mM of POPC in water was chlorinated, after which the solvent was changed to a mixture of d4-MeOH and d1-chloroform (1:5, 600 µL) and analyzed by 1H NMR. An untreated control sample of POPC was prepared and analyzed similarly. The results of the analysis were, however, inconclusive due to the complexity of the chlorinated sample. For easier interpretation, the various chlorinated molecular species would need to be isolated, which is planned for future experiments.
POPC chlorinated in chloroformSamples prepared by chlorination of POPC dissolved in chloroform and untreated control samples were also analyzed with LC-HRMS. The samples were prepared in an attempt to determine whether an aqueous environment is necessary for the production of the novel lipids, as well as to study whether a water-free environment produces additional chlorinated PCs that could give more structural insight into the location of the chlorine substituents in the novel lipids.
No chlorinated lipids were detected in the control sample. Most of the chlorinated lipids observed in the samples chlorinated in water were detected in the chloroform samples as well; however, eight additional chlorinated PCs were also identified (Table 1, Fig. 4). The isotopic patterns and fragmentation measured by LC-HRMS and LC–MS/HRMS indicated that four of the eight additional compounds were peroxy-diphospholipids (PC F, G, H, and I) (Fig. S15–S18 in the SM). The other four had chlorine atoms attached to their glycerol backbone (PC K and L), or to both the glycerol backbone and the acyl chain (PC E and J). They produce fragments containing the glycerol backbone with one or both acyl chains, which support the proposed chlorination site being located in the glycerol backbone (selected structures in Fig. 3). For example PC E and J produce fragments corresponding to ions resulting from the loss of the saturated acyl chain and the loss of the phosphocholine (m/z 681.41777, 626.25415, 715.37880, and 660.21518) (selected structures in Fig. 3). These fragments show that the chlorine atoms must reside in the oleoyl-chain and/or the glycerol backbone. Since the double bonds in both molecules have been saturated with chlorine, the only logical location for the remaining chlorine atoms is the glycerol backbone. The corresponding fragments (m/z 645.44109, 590.27747, 643.42544, and 624.23850) and the retention time of PC K and L support the conclusion. The fragmentation patterns and isotopic patterns of the analytes can be found in Tables S10, S11, S13–S16, S18, and S19 and Fig. S14, S19–21, and S35–S42 in the SM. The proposed structures for all chlorinated PCs are presented in Fig. 4, and their masses are listed in Table 1. The measured accurate masses of all chlorinated PCs did not differ more than ± 2.5 ppm from the theoretical value.
Chlorination of POPC dissolved in chloroform resulted in the production of individual lipids with higher degrees of chlorination than obtained in the chlorination of an aqueous POPC-sample. In water, the differences in hydrophobicity of the headgroup and the alkyl chains cause the lipids to arrange in bilayer vesicles, especially when energy is introduced into the system e.g. through sonication. Although chlorine is a small molecule, the ordered structure of the bilayer vesicles may limit access to the unsaturated sites, and electrostatic and other physical interactions with headgroups nearby may restrict reactivity at the glycerol backbone. In addition, the reaction of chlorine with water (producing HOCl and HCl [29]) competes with the chlorination of the lipids. In chloroform, the lipids are ordered as inverted micelles, in which the acyl chains are oriented towards the solvent. There are fewer competing reactions with the solvent, resulting in more polychlorinated mono- and dimeric-PCs. Due to the differing behavior of lipids and reactions of chlorine in chloroform and in aqueous systems, the chloroform experiments are poor simulations of chlorination reactions in biological systems. Therefore, the suitability of the polychlorinated PCs identified in the chloroform experiments as biomarkers should be critically evaluated. Some polychlorinated PCs may also be formed in tissues (aqueous systems) from chlorine produced in the spontaneous degradation of externally introduced or metabolically produced HOCl. Dichlorination of unsaturated sites through natural metabolism has been demonstrated by the detection of dichlorinated cholesterol in tissue [4, 14]. However, when chlorine is used as a CW, the concentrations are very high. Therefore, it is feasible that polychlorinated lipids, such as the PCs identified in the chloroform experiments, would be more prominent in biomedical samples of chlorine attack victims, than in samples from patients suffering from immunological stress conditions.
Chromatograms of the POPC sample chlorinated in chloroform and analyzed with the different separation methods (C18 and HILIC) are presented in Fig. S1 and Fig. S2 in the SM. From the chromatograms, one can note that the chlorinated PCs elute in three groups on both stationary phases. The first eluting group consists of the PCs chlorinated only at the glycerol backbone (PC A, K, and L). The POPCs chlorinated at the acyl chain double bond (PC D, E, and J) are eluted next. With the HILIC column, PC HOCl elutes in the second group. With the C18 column, the increased interactions with the mobile phase cause the PC HOCl to elute before the first group of chlorinated lipids. The peroxy-diphosphocholines (PC C, F, G, H, and I) are eluted last on both stationary phases. This elution behavior taking place in groups is useful when determining the possible location of the chlorine atom(s) in the lipids. Multiple chromatographic peaks can be detected for most of the chlorinated PCs. This may have resulted from the analytes having several isomers. No conclusions from the structural differences could be drawn from the mass-spectrometric fragmentation patterns of the isomers.
Mechanism of dimerizationAs can be seen from the structures presented in Fig. 4, all chlorinated peroxy-diphosphocholines (PC B, C, F, G, H, and I) have at least one chlorine substituent in the glycerol backbone. Our hypothesis is that the dimerization is initiated by the formation of a PC chlorinated at the glycerol, e.g., PC A. The chlorine substituent changes the three-dimensional structure and electron distribution in the headgroup, allowing dimerization with other lipids in its proximity. After dimerization, the lipids can be further chlorinated at the glycerol and remaining unsaturated sites in the acyl chains. The literature describes the production of peroxy-diphosphates by electrolysis of phosphate solutions containing added halide or pseudohalide anions [30]. However, the exact mechanism of the chlorination and subsequent dimerization of phospholipids cannot be determined from our data. Further experiments are needed to elucidate the mechanisms involved.
Alkaline and enzymatic degradation of chlorinated POPCIn an attempt to verify the position of the chlorine substituent in PC A, chlorinated aqueous POPC samples were treated with mild alkaline hydrolysis or with PLC and analyzed with positive full-scan LC–MS and positive precursor ion scanning of m/z 184 (LC–MS/MS). Positive precursor ion scanning of m/z 184 is a PC-specific MS/MS-scanning mode [31]. The rationale was to investigate whether alkaline or enzymatic degradation would result in degradation products, that would confirm the position of the chlorine in PC A (e.g. a chlorinated glycerophosphocholine or a chlorinated diacylglycerol (DAG)). Mild alkaline hydrolysis degrades PCs to fatty acids and a glycerophosphocholine [32]. The free fatty acids were detected in alkaline-treated chlorinated POPC samples; however, the chlorinated glycerophosphocholine was not detected (data not shown). No chlorinated species were found, which might be a result of the alkaline conditions causing elimination of the chlorine substituent.
PLC is an enzyme that is capable of releasing the phosphocholine headgroup of PCs, yielding a corresponding diacylglycerol lipid [33, 34]. PLC-treatment was able to liberate phosphocholine from POPC, PC HOCl, and PC D, i.e., monomeric PCs without a chlorine substituent in the glycerol backbone (Fig. 1 and Fig. 4). The liberation was evidenced by the loss of respective PC signals (Fig. S50 in the SM) and the appearance of the corresponding DAG in the organic phase (Fig. S52 in the SM). Interestingly, PC A, which putatively contains a chlorine substituent in the glycerol backbone, was seemingly resistant to PLC degradation, as evidenced by apparently no loss of signal intensity between PLC-treated and -untreated samples (Fig. S50 and Fig. S51 in the SM). Masses corresponding to a DAG released from PC A could not be observed in any of the PLC-treated samples; however, the signal may have been below the limit of detection of the instrument.
The signals of PC B and PC C also disappeared after PLC-treatment (Fig. S53 in SM), but no logical degradation products could be identified for these compounds. It is unlikely that the PLC-enzyme was able to cleave the dimers (PC B and C) back into individual lipids, because the signal for PC A in the sample did not increase compared to that of an untreated sample (Fig. S51 in the SM). Nevertheless, the resistance of PC A to degradation by PLC further suggests that the chlorine is located in a position close to the lipid headgroup, possibly hindering PLC from accommodating the lipid headgroup at its active site. The resistance to degradation could also have biological implications. Hindering cellular PC catabolism could affect the generation of lipid signaling molecules (e.g., phosphatidic acid and DAG) that are involved in a range of cellular processes [35].
Chlorinated PSPC and POPETo determine whether other phospholipids in addition to POPC react similarly to chlorination, PSPC and POPE were evaluated. They were dissolved in chloroform or sonicated in water, and subsequently chlorinated and analyzed with LC-HRMS and LC–MS/HRMS, using HILIC separation. Chlorinated lipids were not detected in the control samples that had not been exposed to chlorine gas. Interestingly, no chlorinated PSPC-related lipids were found in any of the samples. The saturated acyl chains of PSPC may have formed more rigid membrane structures, sterically obstructing chlorine from reaching the glycerol backbone. The presence of unsaturated acyl chains (such as the oleoyl in POPC) is seemingly required for sufficient micellar and vesicle bilayer fluidity, and subsequent chlorination of the glycerol backbone. Further experiments are needed to verify this assumption.
Seven chlorinated PEs were detected in the chlorine-exposed POPE samples (Table 1, Fig. 5). The isotopic patterns and fragmentation patterns of the chlorinated PEs are presented in Fig.S23–S29 and S43–S48 and Tables S20–S28 in the SM. The seven chlorinated PEs included the chlorohydrin (PE HOCl), the chlorine saturated PE (PE A), and five PEs chlorinated at the glycerol backbone (PE B–F). The proposed structures of PE A–F are presented in Fig. 5. All seven chlorinated PEs were detected in the POPE sample chlorinated in chloroform. PE F was not produced, and only small amounts of PE D was formed when POPE was chlorinated in water. This difference is presumably caused by the solvent effects described in the “POPC chlorinated in chloroform” section. The accurate masses of some dimeric chlorinated PEs were detected, but their low quantity did not allow reliable identification using isotopic patterns and LC–MS/HRMS analysis. The experiment with POPE showed that other unsaturated phospholipid classes can also be chlorinated in the glycerol backbone, but the reactivity of different lipid classes varies.
Fig. 5
Proposed structures of PE HOCl and PE (A–F), as detected by LC-HRMS. The chlorine(s) may be located in any of the carbons in the glycerol backbone
The chromatograms of the chlorinated POPE samples are presented in Fig. S5–S7 in the SM. The retention of the chlorinated PEs follows the trends observed for the chlorinated PCs (section “POPC vesicles chlorinated in water”). However, there is a notable difference in the retention of the backbone-chlorinated PEs (PE D–F) and the PCs (PC A, K, and L) using HILIC separation (Fig. S2, S4, S6, and S7 in the SM). The backbone-chlorinated PCs eluted near 7 min, while their PE analogues eluted at 1.6 min (PE D) and 0.6 min (PE E and F). This demonstrates that the addition of chlorine atoms to the backbone of POPE reduces the polarity of the resulting PE more compared to the corresponding PCs. The methyl substituents on the amine make the headgroup of PCs bulkier, which may hinder their interaction with the chlorine substituents in the glycerol backbone and the 3D-structure of the chlorinated PCs. The much smaller unsubstituted amine in PEs may allow the headgroup to interact more efficiently with the chlorine atoms in the glycerol backbone, making the molecule less polar (almost non-polar in the case of PE E and F).
Chlorinated pig lung sampleA sample of pig lung was chlorinated in vitro to determine whether the novel chlorinated lipids could be used as biomarkers for chlorine gas exposure. Lung tissue was selected as a chlorination target because inhalation is one of the main exposure routes for chlorine gas. The extracts of the chlorinated and untreated pig lung samples were analyzed using LC-HRMS for chlorinated PCs. Since both POPC and POPE were naturally present in the lung samples, we searched for the accurate mass of PC HOCl, PC A–L, PE HOCl, and PE A–F in the total ion chromatograms (TICs) (Table 1). The TICs were also evaluated for the presence of other chlorinated PCs. The chlorinated PCs were distinguished from the unmodified PCs based on the accurate masses and their isotopic patterns; the patterns had to indicate that 37Cl is present in the molecule, and the monoisotopic peak and the 13C-peak of the dimeric compounds had to be approximately 0.5 mass units apart.
No chlorinated lipids were detected in the untreated control sample. As expected based on previous studies [14,15,16,17,18], PC HOCl and PC D were found in the chlorinated lung sample (Tables S2 and S9 in the SM). PC A and three of the novel chlorinated lipids (PC C, F, and J) were also found in the chlorinated lung sample. The fragmentation patterns of the analytes measured with LC–MS/HRMS matched with the fragmentation of the reference chemicals (Tables S4, S7, S12, and S17 in the SM), suggesting that the detected analytes are chlorinated PCs derived from POPC or structural isomers of POPC. Other chlorine-containing doubly charged PCs (based on the isotopic pattern) were also observed in the chlorinated lung sample at 12.4–12.8 min, which is a typical retention time for chlorinated dimeric PCs using HILIC separation. This indicates that chlorine exposure causes dimerization of other PC molecular species as well. Logically, the structures of the chlorinated mono-PCs and peroxy-diphospholipids formed are dependent on the lipids present in the tissue upon chlorine exposure. Identification of the exact structure of all the various mono- and dimeric chlorinated lipids detected in the lung sample is difficult, due to the vast number of structurally distinct PCs (and other lipids) originally present in the sample.
We propose that all chlorinated lipids identified by us here could potentially be used as biomarkers for chlorine gas exposure, alongside with the previously reported chlorohydrins and dichlorides. However, the compounds with more than two chlorine substituents may only be found in victims exposed to high levels of chlorine gas. The chlorinated derivatives of POPC may be particularly suitable as biomarkers of chlorine gas exposure, due to the ubiquitous nature of this lipid. Nevertheless, since the overall lipid composition is tissue-dependent, other suitable lipid alternatives should be explored. Further effort should be directed towards determining whether glycerol-chlorinated lipids and chlorinated peroxy-diphospholipids can be detected in biomedical samples (e.g., BALF and NLF) in concentrations that allow verification of chlorine gas exposure. The risks of false positives due to natural chlorination by metabolically produced HOCl should also be assessed.
Comments (0)