
Remember me
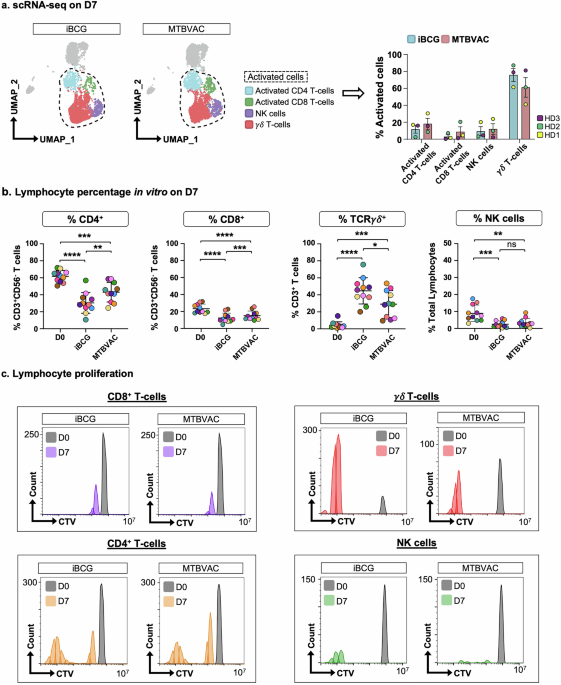

PBMC from three healthy donors were incubated with either MTBVAC or BCG-Moreau RJ strain (iBCG), without exogenous addition of cytokines and, 7 days later, cells were recovered for either single cell RNA sequencing (scRNA-seq) or FACS analyses. A first scRNA-seq analysis of this dataset was done including paired data obtained in parallel for iBCG22 and MTBVAC (Quality control in Supplementary Fig. 1). Large proportions of activated lymphocyte populations, mainly CD4 and γδ T-cells, were identified 7 days after exposure to both vaccines (Fig. 1a, Supplementary Fig. 2), and these data were confirmed by flow cytometry (Fig. 1b, Supplementary Fig. 3) and in proliferation experiments (Fig. 1c). Initial inspection indicated a similar lymphocyte population distribution with both vaccines. Interestingly, compared to iBCG, more CD4 T-cells and less γδ T-cells expanded with MTBVAC. Although CD8 T-cells and NK cells were minority populations, the scRNA-seq data indicated that, together with CD4 and γδ T-cells, they had clear cytotoxic and effector potential, as shown by expression of GZMB, TNFSF10, FASLG, IFNG (Supplementary Fig. 2).
Fig. 1: MTBVAC selectively enhances proliferation of CD4 and γδ T-cells.
a PBMC clusters. PBMC from 3 healthy donors were incubated with either iBCG or MTBVAC for 7 days. scRNA-seq analysis allowed identification of 4 clusters of activated lymphocytes, highlighted in the UMAP plot (left). The frequency of these clusters within the total of iBCG or MTBVAC-activated cells is shown in bar plots (right); mean ± SD are depicted. Dots represent each healthy donor in a different colour. b Lymphocyte activation by FACS. PBMC from 11 healthy donors, depicted in different colours, were incubated with either iBCG or MTBVAC. After a week, cells were recovered, and the different subsets analysed by flow cytometry. Scatter plots show mean ± SD. Statistical analysis was done by paired sample t-test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001) comparing % of activated cells with each vaccine against % on basal condition, D0, and between vaccines. c Activated lymphocytes proliferation. Profiles of lymphocyte subsets after one week of co-culture were analysed by flow cytometry. Data from a representative healthy donor are shown.
Although in the initial scRNA-seq analysis cell groups converged in the same cluster based on sharing a certain number of genes, this does not mean they are 100% identical23. Indeed, when scRNA-seq data of MTBVAC-stimulated PBMC were studied individually in depth to gain detailed insight into this immune response (Fig. 2; quality criteria Supplementary Fig. 1), 12 clusters defined by differential gene expression were identified and annotated (Fig. 2a, b, Supplementary Fig. 4). MTBVAC-stimulated PBMC yielded 3 clusters of CD4 T-cells (clusters 2, 3, 4) and 3 clusters within a large γδ T-cell population (clusters 1, 6, 8).
Fig. 2: scRNA-seq analysis of MTBVAC-stimulated PBMC.
a PBMC clusters. The MTBVAC-primed dataset from 3 healthy donors was analysed and clusters annotated using the FindClusters function from Seurat R package (see Materials and Methods). UMAP plot represents the 12 clusters identified. Selected clusters are grouped: CD4 (2, 3, 4),  ; CD8 (5, 10),
; CD8 (5, 10),  ; γδ T-cells (1, 6, 8),
; γδ T-cells (1, 6, 8),  . b Cluster characterization. Dot plot represents average expression (colour graph) and % of cells (dot size) for markers (x-axis) defining each cluster (y-axis).
. b Cluster characterization. Dot plot represents average expression (colour graph) and % of cells (dot size) for markers (x-axis) defining each cluster (y-axis).
Cytokine-producing CD4 T-cells are thought to play a key role in the control of Mtb infection. Thus, differential gene expression of the three distinct CD4 T-cell clusters (C2: naïve, C3: memory, C4: activated) that were present following MTBVAC co-culture was analysed (Fig. 2). As previously described for other mycobacteria, the phenotype of the activated CD4 population (C4) expanded with MTBVAC corresponds mainly with Th1, with increased RNA transcripts for CXCR3, IFNG and lytic granule components, as well as CCL5 (Fig. 3a). Further analysis of the activated Th1 cluster of CD4 T-cells revealed 2 clear subpopulations (Fig. 3b–d): SC1, enriched in IL7R and AQP3 (aquaporin-3), consistent with a memory, less activated subpopulation; while SC2 had markers consistent with a potent Th1 activation, displaying a strong expression of molecules like IFNG, CSF2, IL26 or TNF, but not IL17A.
Fig. 3: Characterisation of MTBVAC-stimulated CD4 T-cells.
a Violin plots. Violin plots show expression level of signature markers for the CD4 clusters (2, 3, 4) (x-axis) identified in Fig. 2. b–d Activated CD4 T-cell analysis. UMAP plot of cluster 4 from Fig. 2 (b) identified two subclusters ( ). The 10 most differentially expressed genes of SC1 and SC2 are shown in a heatmap (c). Feature plots highlight the expression of IFNG, CSF2, IL26, TNF, IL21 and IL17A within activated CD4 T-cells (d). e CD4 T-cell differences after exposure to vaccines. iBCG and MTBVAC-stimulated CD4 T-cells were analysed by differential marker expression (x-axis) in a dot plot. The average expression level and percentage of cells expressing a particular marker is shown using colour density and dot size respectively. f, g Activation markers by FACS. Representative plots show HLA-DR, CD62L, CD25 and CXCR3 markers within CD3+ TCRγδneg CD4 T-cells from PBMC on D0 (f) and on PBMC one week after incubation with MTBVAC (g). Gating strategies are shown for both resting and activated states. For activated PBMC (g), the frequency of selected populations is plotted in either bar graphs or scatter plots (n = 5; mean ± SD). For the CD4hi CD62Lneg HLA-DR+ CD25+ subpopulation, a scatter plot shows the frequency against basal state, D0, and between vaccines compared by a by paired sample t-test (ns: p > 0.05) for the 5 healthy donors, depicted in different colours.
). The 10 most differentially expressed genes of SC1 and SC2 are shown in a heatmap (c). Feature plots highlight the expression of IFNG, CSF2, IL26, TNF, IL21 and IL17A within activated CD4 T-cells (d). e CD4 T-cell differences after exposure to vaccines. iBCG and MTBVAC-stimulated CD4 T-cells were analysed by differential marker expression (x-axis) in a dot plot. The average expression level and percentage of cells expressing a particular marker is shown using colour density and dot size respectively. f, g Activation markers by FACS. Representative plots show HLA-DR, CD62L, CD25 and CXCR3 markers within CD3+ TCRγδneg CD4 T-cells from PBMC on D0 (f) and on PBMC one week after incubation with MTBVAC (g). Gating strategies are shown for both resting and activated states. For activated PBMC (g), the frequency of selected populations is plotted in either bar graphs or scatter plots (n = 5; mean ± SD). For the CD4hi CD62Lneg HLA-DR+ CD25+ subpopulation, a scatter plot shows the frequency against basal state, D0, and between vaccines compared by a by paired sample t-test (ns: p > 0.05) for the 5 healthy donors, depicted in different colours.
Since there were similarities in the cell populations in MTBVAC and iBCG-stimulated PBMC, detailed analyses of the differences between the effector populations were carried out. For CD4 T-cells, the main differences between the two vaccines occurred in MHC class II gene expression (Fig. 3e): HLA-DP, DQ, DR and DM were less expressed after MTBVAC stimulation, while SELL [L-selectin (CD62L)] transcripts were only found in CD4 T cells activated by culture with MTBVAC, but not BCG. These findings could reflect differences in CD4 T cell activation after exposure to either MTBVAC or iBCG.
The phenotype of activated CD4 T-cells was confirmed by flow cytometry (Fig. 3f, g; Supplementary Fig. 5). As expected, the CD25 activated subset was negative for CD62L and expressed HLA-DR, while the CD62L-positive population was negative for HLA-DR and CD25. Moreover, a clear increase of CXCR3 was observed in activated CD4 T-cells after stimulation with both vaccines, iBCG and MTBVAC.
Altogether, MTBVAC stimulation activated a CD4 T-cell population with strong expression of MHC-II, presumably proliferating after antigen encounter, and homing molecules like CXCR3.
MTBVAC stimulates CD16+GZMBhi cytolytic γδ T-cells expressing CD62Lγδ T-cells have been proposed to play important roles in Mtb control18. In fact, three distinct γδ T-cell clusters (C1, C6, C8) (Fig. 2a) were observed to proliferate actively after MTBVAC stimulation. scRNA-seq analysis of the γδ T-cell population obtained after a 7-day incubation of PBMC with MTBVAC showed that these cells were mainly Vγ9Vδ2 (TRDV2, TRGV9) (Fig. 2b). Further in-depth analyses revealed 7 subclusters (Fig. 4a, Supplementary Fig. 6a), many of them expressing CD27 (Fig. 4b). Moreover, a marked expression of IFN-γ was found, especially in SC3, which represents an active subpopulation (IL2RA+IFNG+) (Fig. 4c). CD27+ IFNγ+ γδ T-cells have been described as pro-inflammatory with anti-tumour capacity, as opposed to suppressor CD27- IL17A+ γδ T-cells16. As expected from the CD27+ IFNγ+ phenotype, MTBVAC-primed γδ T-cells did not express IL17A. In line with this, both TB vaccines, BCG and MTBVAC, stimulated γδ T-cells to express high levels of cytotoxic molecules like GZMM, GZMB and GNLY, proinflammatory chemokines like CCL5 (RANTES), and receptors like CXCR3 (Fig. 4d). Despite these similarities, transcriptomic analysis of γδ T-cells also identified some differences between MTBVAC and BCG-activated PBMC: MTBVAC stimulation led to more cells expressing molecules like HAVCR2 (TIM3) and TNFRSF9 (4-1BB), albeit at low levels. These molecules have been associated with either T-cell activation or exhaustion in different contexts24,25,26. So, while a low TIM3 expression could denote some level of exhaustion, proinflammatory and activation markers would argue against this possibility for γδ T-cells. IFNG and GZMB were expressed in higher levels in MTBVAC while GZMM was slightly lower than in BCG-primed cells. In contrast, BCG-stimulated cultures showed a higher expression of genes related with type I IFN responses (e.g. ISG20, IFITM1). These findings indicate that activation of γδ T lymphocytes after stimulation with non-pathogenic mycobacteria can initiate a proinflammatory response and participate in homeostasis regulating infiltration of CD8 T-cells27,28.
Fig. 4: Characterisation of MTBVAC-stimulated γδ T-cells.
a–f scRNA-seq. a UMAP plot represents γδ T-cell subclusters from the 3 healthy donors of Fig. 2. b Violin plot showing expression of CD27 for each γδ T-cell subcluster. c Feature plot of IFNG and IL2RA markers in γδ T-cells. d Dot plot of markers (x-axis) discriminating γδ T-cells from iBCG and MTBVAC experiments. The average expression level and percentage of cells expressing a particular marker are shown using colour density and dot size respectively. e Violin plots comparing the expression of FCGR3A in γδ T-cells from MTBVAC and iBCG cultures. f FCGR3A expression in γδ T-cells from MTBVAC cultures: UMAP highlighting γδ T-cells with positive values of FCGR3A (left) and volcano plot of differentially expressed genes between γδ T-cells expressing (POS) or not expressing (NEG) FCGR3A (right). Genes with a Log2FC > 0.58 and a Bonferroni adjusted p-value < 0.05 are marked in  . g γδ T-cell phenotype by FACS. PBMC from 5 healthy donors were incubated with either iBCG or MTBVAC for a week and cells were analysed by flow cytometry. For each marker, representative histograms and MFI scatter plots (mean ± SD), with each donor represented in different colour, are shown. Statistical analysis of % of expression against expression on basal condition, D0, and between vaccines was done by paired sample t-test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
. g γδ T-cell phenotype by FACS. PBMC from 5 healthy donors were incubated with either iBCG or MTBVAC for a week and cells were analysed by flow cytometry. For each marker, representative histograms and MFI scatter plots (mean ± SD), with each donor represented in different colour, are shown. Statistical analysis of % of expression against expression on basal condition, D0, and between vaccines was done by paired sample t-test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
Since individuals from TB endemic regions that control the infection have an expanded population of CD8+ γδ T-cells expressing the low affinity IgG receptor FcγRIII (CD16) that may help to control bacteria18, we also evaluated these markers. FCGR3A was found in 21.2% of MTBVAC-primed γδ T-cells (Fig. 4e, f, Supplementary Fig. 6b), which is remarkable since, usually, this transcript is expressed at low levels in this subset. γδ T-cells expressing FCGR3A contained more markers of a cytotoxic profile, such as GNLY, GZMB, CX3CR1 (Fig. 4f). In contrast, iBCG priming resulted in a lower percentage of γδ T-cells expressing FCGR3A (14.4% in iBCG). Flow cytometry of γδ T-cells expanded after MTBVAC co-incubation confirmed that nearly 40% of γδ T-cells expressed significantly more CD16 (Fig. 4g) than those activated after BCG (20%). A small CD8+ γδ T-cell population (mainly CD8α+) was also identified after incubating PBMC with either iBCG or MTBVAC vaccines (Supplementary Fig. 6c–e).
Besides expression of cytotoxic and proinflammatory molecules, the entire γδ T-cell population was activated, as cells had upregulated expression of CD25, HLA-DR and CXCR3 by flow cytometry (Fig. 4g). These γδ T-cells also expressed several NK receptors (Supplementary Fig. 4a), like NKG2D, but no PD1 expression was found (Fig. 4g). It is also intriguing that secondary lymphoid homing molecules have very different expression levels: while CD62L was increased in the whole γδ T-cell population, CCR7 was almost absent (Fig. 2b). Interestingly, in a bovine model, γδ T-cell skin egress and migration into lymph nodes was proposed to be mediated by CD62L expression, independently of CCR729.
In aggregate, a differentiated effector population loaded with cytolytic molecules, consistent with the CD16+GZMBhi cytolytic γδ T-cells identified in individuals that controlled Mtb infection18, was also found in MTBVAC-expanded PBMC. These cells can respond through antibody (via CD16) and NKG2D-mediated cytotoxicity and may have the capacity to home to secondary lymphoid organs. However, whether this population can be maintained over time after vaccination is as yet unknown.
After MTBVAC co-culture, NK and CD8 T-cell percentages decrease, but these populations are activatedIn contrast to CD4 and γδ T-cells, the proportion of CD8 T lymphocytes and NK cells clearly decreased after MTBVAC stimulation of PBMC (Fig. 1c). Nevertheless, although the number of CD8 T-cells identified was low, an obvious cluster of cytotoxic CD8 T-cells (C10) could be distinguished from another of naïve CD8 T-cells (C5) (Fig. 2). High levels of GZMB and CCL4 defined the cytotoxic CD8 T cell cluster (Supplementary Fig. 7a) which also clearly expressed the transcription factors ZNF683, a marker of tissue resident memory, as well as ZEB2, the chemokine receptor CX3CR1, the integrin ITGB1 and the innate molecules KLRD1 and KLRG1, the latter associated with terminal differentiation of effector cells30. These cells were similar to the ones identified in iBCG cultures, but expressed more DDX5, further confirming an effector activation phenotype31, and less ITGA4 (CD49d), present in virtual memory cells (Supplementary Fig. 7b). CD38 was also low in MTBVAC-primed cultures. Although expression of this marker usually occurs after T cell activation, a CD38/CD203/CD73 axis can also diminish T-cell functions32,33. In fact, other data indicate that low CD38 expression discriminates reprogrammable T-cells from PD1hi dysfunctional subsets34. So, MTBVAC-stimulated PBMC contain activated cytotoxic T-cells that are less likely to be exhausted.
Taken together, these data indicate that a small subset of non-exhausted effector CD8 T-cells can be clearly defined in cultures responding to MTBVAC stimulation.
Two subclusters of NK cells could also be identified in scRNA-seq, both with cytotoxic features and expressing molecules consistent with a CD56dim phenotype (Fig. 5a, b). However, SC2, expressing both IL2RA and IFNG (Supplementary Fig. 7c), represents a more active subset than SC1. SC2 also expressed FASLG, TNFRSF18, CSF1 and IL21R, and so are NK cells with great potential to kill target cells (Fig. 5c). Comparison between MTBVAC and BCG-primed NK cells revealed only small differences (Fig. 5d). Notably, although GZMB was strongly transcribed in NK cells from both cultures, its expression was even stronger after MTBVAC-stimulation. Taken together, these features of NK cells indicate that they might potently kill target cells.
Fig. 5: Characterisation of MTBVAC-stimulated NK cells by scRNA-seq.
Analysis of NK cell subpopulations from MTBVAC-primed PBMC identified in 3 healthy donors in Fig. 2. a UMAP. Plot shows two NK cell subclusters ( ). b Volcano plot. Genes differentially expressed between NK cell subclusters
). b Volcano plot. Genes differentially expressed between NK cell subclusters  and
and  are highlighted, marking in
are highlighted, marking in  genes with a Log2FC > 0.58 and a Bonferroni adjusted p-value < 0.05. c Matrix plot. Expression of markers within each NK cell subcluster are indicated according to colour scale. Markers differentially detected between both subclusters are highlighted with an asterisk. d Dot plot. Different markers (x-axis) in NK cells from iBCG and MTBVAC experiments. The average expression level and percentage of cells expressing a particular marker are shown using colour density and dot size respectively.
genes with a Log2FC > 0.58 and a Bonferroni adjusted p-value < 0.05. c Matrix plot. Expression of markers within each NK cell subcluster are indicated according to colour scale. Markers differentially detected between both subclusters are highlighted with an asterisk. d Dot plot. Different markers (x-axis) in NK cells from iBCG and MTBVAC experiments. The average expression level and percentage of cells expressing a particular marker are shown using colour density and dot size respectively.
The range of cytokines produced by PBMC, after a 7-day co-culture with MTBVAC, were studied by analysing mRNA transcripts in the different cell populations identified in scRNA-seq, and these experiments were complemented by Luminex analyses of a panel of effector and suppressor cytokines secreted into tissue culture supernatants of MTBVAC- and iBCG-stimulated PBMC (Fig. 6).
Fig. 6: Cytokine profile after 7-day exposure to vaccines.
PBMC from 6 healthy donors (depicted in different colours) were incubated with either iBCG ( ) or MTBVAC (
) or MTBVAC ( ). After 5, 7 and 9 days in culture, cell supernatants were recovered for cytokine quantification by Luminex. Scatter plots show mean ± SD for each condition. Limits of detection are indicated in grey. Statistical analysis of cytokine concentration in activated cultures compared to the untreated condition (
). After 5, 7 and 9 days in culture, cell supernatants were recovered for cytokine quantification by Luminex. Scatter plots show mean ± SD for each condition. Limits of detection are indicated in grey. Statistical analysis of cytokine concentration in activated cultures compared to the untreated condition ( ) and between vaccines on the same day was done by paired sample t-test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
) and between vaccines on the same day was done by paired sample t-test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
Consistent with the observation that MTBVAC stimulation led to expansion of mainly IFN-γ-producing lymphocyte populations, the presence of this cytokine was clearly detected in supernatants, while IL-2 and IL-4 were almost undetectable. TNF-α and IL-6 were also produced by MTBVAC-activated PBMC (Fig. 6a), while IL-17A and IL-17F were only detected in extremely low concentrations. These data confirmed the scRNA-seq analyses, in which the secretion of each cytokine can be associated to the producing cell subcluster (Supplementary Fig. 8a). Feature-plots of the cytokines secreted by activated CD4 T-cells, revealed high levels of IFNG, CSF2 (GM-CSF) and IL26 transcription (Fig. 3d). Interestingly, IL26 has been described to have anti-microbial function and bind mycobacteria derived lipoarabinomannan (LAM)35. On the other hand, TNF and IL21 were only expressed in low amounts by CD4 cells and IL17A was virtually absent. As mentioned before, γδ T-cells also transcribed high levels of IFNG, TNF and IL15 (Supplementary Fig. 8b).
Taken together these data confirm the secretion of cytokines expected from a majority Th1 CD4 T-cell population, such as IFN-γ and TNF-α. However, the concentration of these factors was enhanced by the contribution of a specialized effector γδ T-cell population.
BCG has been used for decades as therapeutic agent against bladder cancer and NK cells activated with BCG are very efficient killing tumours via NKG2D36. Therefore, it was of interest to test whether MTBVAC could have some role as a cancer therapeutic. We confirmed that MTBVAC-primed NK cells increased NKG2D cell surface expression (Supplementary Fig. 9a). Killing assays using MTBVAC-primed PBMC, as effectors, against several bladder cancer cell lines as target cells (Supplementary Fig. 9b–d) revealed that, although the levels of tumour target lysis were in general low, they could be related to the percentage of NK cells present in the cultures at day 7. Degranulation experiments showed that NK cells (Fig. 7a), and to a lesser extent γδ T-cells (Supplementary Fig. 10a), were the main subpopulation degranulating against K562 control target cells. However, degranulation was generally weak against bladder cell lines. On the other hand, intracellular staining at day 7 showed that NK cells released low or no IFN-γ and TNF-α after recognition of K562 and bladder cancer cells (Fig. 7b, c; Supplementary Fig. 10b, c). These experiments indicate that BCG stimulates anti-tumour NK cells slightly more efficiently than MTBVAC.
Fig. 7: Effector function of stimulated lymphocytes against bladder cancer targets.
PBMC from 5 healthy donors were incubated with iBCG or MTBVAC. After a week in culture, cells were recovered and used as effector cells against solid tumour bladder cancer cells: T24, J82, and RT112 or the positive control NK cell target K562, as indicated. a Degranulation. Activated PBMC were tested as effector cells (1:2 E:T ratio) and degranulation measured as surface LAMP-1 (CD107a). Scatter plots show mean ± SD for n = 5 (each healthy donor represented in a different colour). b, c Intracellular IFN-γ and TNF-α production by FACS. Dot line shows mean value on D7 against no target. Statistical analysis compared to basal expression on D0, was done by paired sample t-test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
Comments (0)