
Remember me
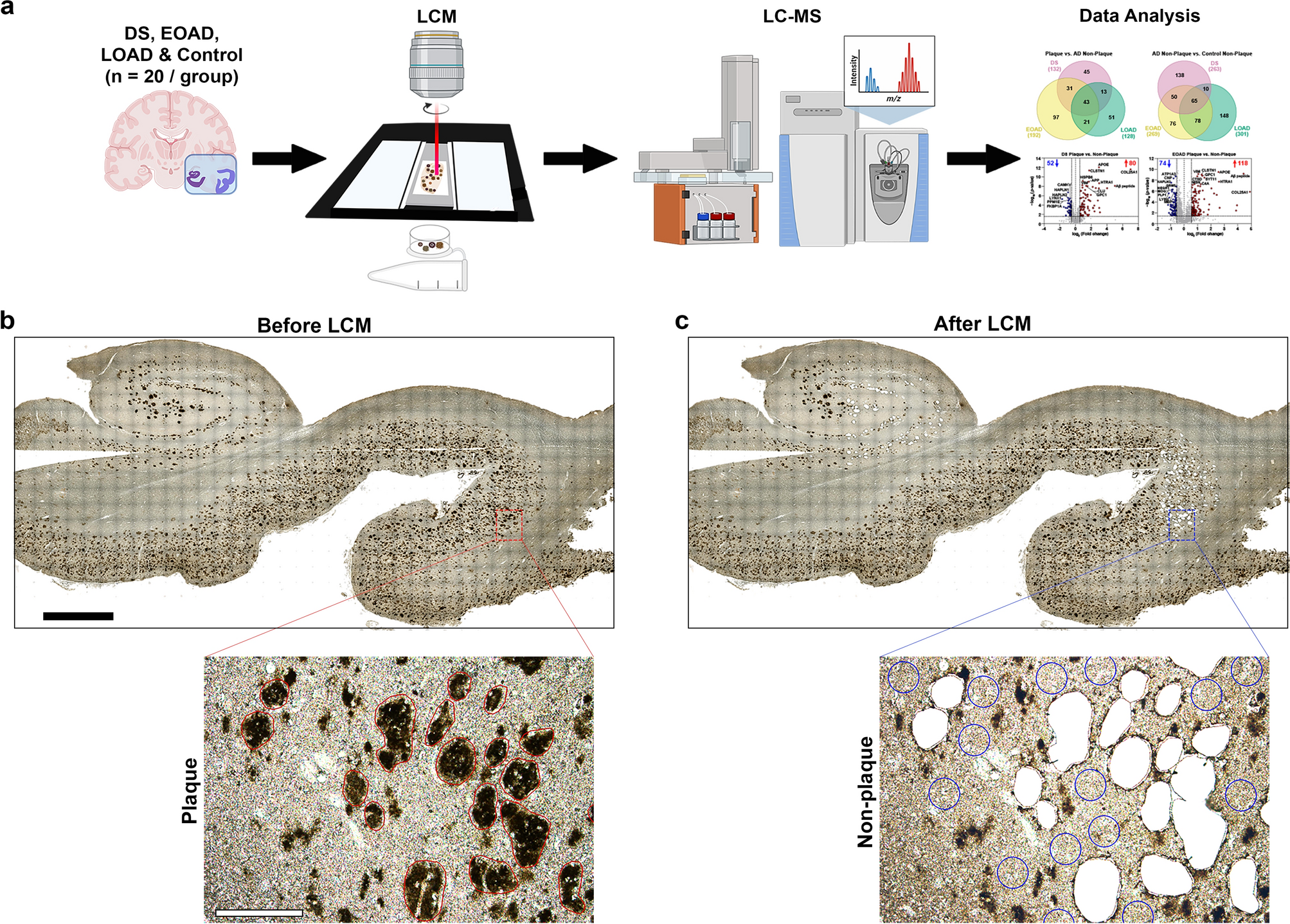

To elucidate the alterations of microglia within the hippocampus of AD patients, we performed IHC analysis of the microglial morphological marker, ionised calcium-binding adaptor molecule 1 (Iba1), in a cohort of CTLs and AD post-mortem FFPE samples. We identified two distinct types of large Iba1-positive cellular accumulations. The most common were the typical Aβ plaque-associated microglia (PaM), characterised by a high number of microglia clustered around an Aβ core plaque in a rosette-like conformation. Less frequently encountered, but consistently present in our sample collection, were clusters of microglia with a polarised shape reminiscent of a 'soma' and 'tail' configuration, observed predominantly in the PL of the CA1 and CA2 regions of the hippocampus (Fig. 1A, B). Across serial sections of hippocampi from 16 AD cases and 5 age-matched CTLs, we observed that this type of accumulation occurred in the CA region of AD patients at a ratio of approximately 1 to 11 compared to PaM. This accumulation was absent in CTL samples and was not detected in prefrontal, temporal, or entorhinal cortices of either CTL or AD cases.
Fig. 1
PaM and CoM have a distinct morphology and relationship to Aβ plaques in hippocampi of AD patients. A–C Representative images of PaM and CoM in human post-mortem hippocampal AD tissue (case #52). A, B Consecutive FFPE sections stained with Iba1 (brown) and nuclei (haematoxylin, blue). C PFA-fixed section stained against Iba1 (magenta) and nuclei (DRAQ7™, white). Both staining methods reveal typical rosette distribution of PaM (full arrows) and polarised accumulations of microglia termed CoM (empty arrows) in the CA1 PL. The asterisk indicates a blood vessel to highlight that these accumulations are not associated with the vasculature. D, E Confocal overview of Iba1 (magenta) and 4G8 (teal) stainings in AD hippocampi showing the co-localisation between PaM (full arrows) and Aβ plaques (D, case #35) while CoM are spatially distant from Aβ plaques (empty triangles) (E, case #38). (F, G) High-resolution 3D confocal images showing the morphological differences between PaM (F, case #25) and CoM (G, case #35) and their respective interaction with Aβ plaques. H Quantification of average volume and number of nuclei of PaM, CoM and CoM-Rod across our collection of AD and DLB and age-matched CTL samples. Images showing confocal 3D stacks of microglia (Iba1, magenta) and nuclei (DRAQ7™, white) with corresponding 3D segmentations with violin plot of volume (µm3) and nuclei (count) of PaM, CoM and CoM-Rod. I CD68 (brown) expression is increased in AD CA1 (case #52) compared to CTL (case #7) and found in PaM (full arrows) and CoM-like accumulations (empty arrows). Scale bars: A 1mm (low mag), 100µm (high mag); B, C 100 µm D, E 1 mm (low mag), 100 µm (high mag); F, G 20 µm; H 50 µm; I 30 µm
To further characterise the 3D morphological features and differences from PaM, we immunostained PFA-fixed 80 µm hippocampal sections from AD patients for Iba1 and 4G8 (Aβ). Using confocal and STED microscopy, we confirmed the enrichment of this peculiar type of Iba1- accumulation in the CA1 and CA2 PL of AD hippocampi (Fig. 1C) and showed that they were spatially separated from 4G8-positive Aβ plaques (Fig. 1D–G, Supplementary Fig. 1A, B). High and super-resolution microscopy revealed new features of their complexity with Iba1-cells showing tight boundaries and an encapsulating cellular organisation that was clearly distinct from PaM [11, 19] with a size and consistent orientation resembling that of pyramidal neurons (Supplementary Fig. 1, Supplementary Movie 1; Supplementary Movie 2). Co-staining with DRAQ7™ often showed nuclei that appeared to be engulfed by Iba1-positive cells (Supplementary Movie 1, Supplementary Fig. 2). These microglia collectively formed a tight shell around as yet-unidentified structures, resembling the glial embrace (“Gliöse Umklammerung”) or coffin formation (“Totenladenbildung”) around neurons observed in various conditions in the early twentieth century literature [2, 27, 41, 87, 89], that we hereafter referred to as “coffin-like microglia” or CoM (Fig. 1A, C, E, G; Supplementary Fig. 1B, D).
Our high-resolution 3D analysis allowed us to distinguish between two types of CoM. We noticed some CoM formations were predominantly composed of bipolar or rod-shaped microglia, resulting in a thin and elongated structure in 3D, often traversing the PL, while only partially enveloping surrounding cells. Unlike the first CoM described, the other type did not seem to enclose any other structure. We termed them CoM-Rod (Fig. 1H, Supplementary Fig. 3). In addition, we often observed smaller groups of Iba1-positive cells 'clumped' together, located between other brain cells (Supplementary Fig. 3). Our previous findings indicated that morphological alterations of individual microglia in DLB resembled to those observed in AD, but with less severity [22]. Consistent with these observations, we detected PaM, CoM, CoM-Rod, and clumped microglia to a lesser extent in the hippocampi of DLB patients (Supplementary Fig. 1C, D).
We quantified CA1/CA2 PaM and CoM number of cells and their total volume from 3D confocal stacks (40x). We did not observe CoM in age-matched CTL samples (n = 8), however, we found few PaM and CoM-Rod. Using Imaris software, we quantified volumes and number of nuclei contained in 61 3D z-stack images of PaM, CoM and CoM-Rod across age-matched control (n = 1), AD (n = 8) and DLB patients (n = 4) samples (Fig. 1H). PaM presented a mean volume of 19,744 µm3 with an average of 20 nuclei, CoM had a mean volume of 15,776 µm3 and an average of 17 nuclei, and CoM-Rod a mean volume of 8,870µm3 and an average of 9 nuclei (Fig. 1H). Using the Kruskal–Wallis test by ranks, we showed that CoM-Rod are significantly smaller in volume (P < 0.05, P < 0.01 respectively) and in nuclei count (P < 0.01) than CoM and PaM. Even though PaM and CoM are very different in morphology, their volume and nuclei count did not present significant differences.
We then investigated the association of a typical marker of microglia upregulated in disease, cluster of differentiation 68 (CD68) [9, 30, 71]. We first quantified CD68 expression in hippocampal subfields of CTL and AD FFPE samples (n = 7 CTL, 8 AD). CD68 was expressed in microglia-like cells in CTL and AD in all brain regions examined, slightly increased in all subfields but more drastically in the CA1 AD (P < 0.05). We found CD68 associated with PaM and CoM-like structures in the CA hippocampus, showing a shared feature of PaM and CoM (Fig. 1I).
PaM and CoM exhibit distinct pathological and astrocytic microenvironmentsTo elucidate the relationship of PaM and CoM with surrounding cells, tau and α-synuclein pathologies, we investigated the characteristics of their pathological microenvironment. Given the distinct association of PaM and CoM with Aβ plaques, we further investigated their interactions with pTau NFTs and pSyn aggregates in AD [22, 97]. Using high-resolution confocal imaging of immunostained sections for Iba1 and AT8 (pTau), we found that CoM can fully encompass NFT-bearing neurons, enveloping the entire 'flame' shape of the tangle and spreading to surrounding neuronal cells (Fig. 2A, B, Supplementary Movie 3). Using thiazine red (TR), which binds β-pleated sheet structures, we also observed CoM containing TR-positive structures resembling NFTs (Supplementary Fig. 4B, Supplementary Movie 4). Similarly, we found some cells harbouring intracellular granular pSyn inclusions enveloped by CoM (Fig. 2C, D, Supplementary Movie 5). Notably, some CoM lacked AT8 tangles (Supplementary Fig. 4A) or pSyn aggregates.
Fig. 2
CoM and PaM are both associated with tau pathology. A, B CoM (Iba1, magenta) are associated with AT8 positive NFTs (cyan) in the PL of CA1 in AD patients (case #23). High-resolution of the CoM reveals its polarised architecture in 3D (left panels) and enwrapping along the NFT in 2D z-stacks (right panels). C, D CoM (Iba1, magenta) can encompass neuronal cells carrying intraneuronal granular pSyn inclusions (11A5, cyan) in the hippocampus of AD patients (AD, case #35). High-resolution of the CoM shows its polarised architecture in 3D (left panels) and close aggregation around the cell bearing granular pSyn inclusion in 2D z-stacks (right panels). E, F 4-plex chromogenic IHC for Iba1 (DAB, brown), AT8 (purple), 4G8 (yellow) and PS396 (teal) highlighted the accumulation of swollen AT8 + PS396 + (dark blue) structures near plaques. We found neurons with tangles that were AT8 + only, PS396 + only or double positive. AT8 + PS396 + were often surrounded by microglia, sometimes forming aggregates reminiscent of CoM (empty arrows) (cases #30, #48 and #49). G When GFAP staining (teal) is combined with that of 4G8 (yellow) and PS396 (DAB, brown), it reveals the encapsulation of plaque-associated PS396 by PaM astrocytes (case #48). Scale bars: A 200 µm; B 50 µm; C 100 µm; D 20 µm; E–G 30 µm
We next examined the distribution of the P-Ser396 pTau (PS396), a preferred site of GSK-3 β phosphorylation activated by phospho-AKT1 (S473) [81], in combination with the S202 and S205 phosphorylated forms of tau labelled by AT8, 4G8 and Iba1 staining in IHC multiplex chromogenic experiments (n = 4 AD). We found relative heterogeneity of PS396 and AT8 tangles and neurites across samples, with some samples showing a distinct ratio of PS396 to AT8 loading (Fig. 2E, F). We observed single stained neurons, AT8 + or PS396 + , and double stained AT8 + PS396 + . The plaque microenvironment defined by 4G8 staining and Iba1 PaM showed a severe tau pathology, with both in AT8 and PS396 labelled swollen structures and often double-positive (Fig. 2E, F). Groups of microglia reminiscent of CoM were also found surrounding neurons with double-positive tangles (Fig. 2F). To understand the compartmentalisation of neurodegenerative processes around Aβ plaques, we used multiplex chromogenic labelling for PS396 Tau, 4G8 and glial fibrillary acidic protein (GFAP), to label PaM astrocytes that were shown to form nets [11]. We found that astrocytes frequently encapsulate PS396 + structures and Aβ plaques (Fig. 2G).
We then investigated in more detail the relationship of PaM and CoM with surrounding astrocytes, whose morphological changes may indicate a specific interplay with microglia. Using chromogenic multiplex IHC, for Iba1, GFAP and 4G8 (n = 5 CTL, 8 AD), we quantified the number of PaM and PaM-associated astrocytes (PaM astrocytes), those polarised around PaM and forming a net, associated with Aβ plaques (n = 391) in the CA1 and measured the corresponding plaque area (Fig. 3A–C). As expected, more PaM than PaM astrocytes were attached to Aβ plaques per area (Fig. 3B). We found a positive correlation between CA1 plaque area, the number of PaM (r = 0.398, ***) and the number of astrocytes forming a net (r = 0.37, ***) (Fig. 3C) with a similar trend as in the cortex [11]. In total, 26.1% of Aβ plaques were encapsulated by both PaM and PaM astrocytes, 22% of Aβ plaques smaller than 2500 µm2, 53.3% of Aβ plaques larger than 2500 µm2. With this approach, there was no evidence of CoM encapsulation by astrocytes. Using 3D confocal microscopy, we compared the distribution and morphology of the astrocytes surrounding PaM or CoM. Around core Aβ plaques (4G8), hippocampal PaM (Iba1) were often surrounded by an outer sphere of hypertrophic and polarised astrocytes (GFAP, PaM astrocytes), as previously observed in AD cortical brain samples (Fig. 3D), but also in APP mouse models [11]. However, we found that astrocytes surrounding CoM ( CoM astrocytes) mainly exhibited a dysmorphic phenotype, with thin and irregular branches, lacking polarisation towards the CoM and showing lower reactivity, here assessed by GFAP intensity (Fig. 3E, Supplementary Fig. 4B). Some GFAP processes were found invading CoM boundaries suggesting a possible interaction at subcellular level different from that around Aβ plaques (Supplementary Movie 4).
Fig. 3
PaM and CoM show a distinct astrocytic surrounding. A 3-plex chromogenic IHC of Iba1 (DAB, brown), 4G8 (yellow) and GFAP (teal) showing PaM astrocytes forming polarised branches in a net-like structure (case #36). B Quantitative 2D analysis of PaM and PaM-astrocytes attached per Aβ plaques area. C The number of PaM and PaM astrocytes shows a positive correlation with the area of Aβ plaques (respectively, r = 0.398 and r = 0.37, **** P < 0.0001) D 3D confocal stack showing PaM (Iba1, magenta) surrounded by reactive astrocytes (GFAP, green) in direct proximity and forming together a RGN around Aβ plaques (4G8, cyan) (AD case #25). E Astrocytes (GFAP, green) do not form a polarised net around CoM (Iba1, magenta) (AD case #25). Scale bars: A 30µm; D 50µm; E 20µm
Digital spatial profiling of 147 proteins highlights core differences between PaM and CoM and their microenvironmentTo delve deeper into the molecular signatures of PaM and CoM in relation to their surrounding microenvironment in AD, we used the Nanostring GeoMx Digital Spatial Profiler (DSP) platform. We quantified 147 proteins in spatially defined CoM (n = 5) and PaM (n = 7) Iba1-positive areas and their close surrounding in FFPE hippocampal sections from three AD patients (cases #49, #50 and #52; Supplementary Table 1). Utilising fluorescent antibodies for microglia (Iba1), astrocytes (GFAP), Aβ-plaques (MOAB-2) and nucleic acid stain (SYTO™ 13), we accurately localised and identified PaM with astrocyte net and CoM. By manually delineating regions of interest for PaM and CoM in the hippocampal PL from the CA2 and CA1, we selected two AOIs: the Iba1-positive area and its immediate surrounding (Fig. 4A). After negative probes normalisation, only a limited number of DEPs reached significance FDR > 0.1, |Log2 fold change (FC)|> 0.5, padj < 0.05) between PaM and CoM and between their respective surroundings (Fig. 4B, Supplementary Excel 1). Her2 (receptor tyrosine-protein kinase erbB2), complement component 4B (C4B), and protein kinase B (Phospho-AKT1, S473) were enriched in PaM compared to CoM while stimulator of interferon genes (STING) exhibited higher levels in CoM. Granzyme A (GZMA), progesterone receptor (PR), and p53 were enriched in vicinity of PaM vs. CoM. Despite not reaching statistical significance, several proteins indicated potential differences in cellular signatures or microenvironments. Purinergic receptor P2RY12, Iba1, and microglia/immune cell-associated proteins such as arginase 1 (ARG1) or lymphocyte activating 3 (LAG3) were elevated in CoM compared to PaM. Amyloid peptides, amyloid precursor protein (APP), GFAP, CD163, CD44 and apolipoprotein E (ApoE) were found at higher levels in both PaM and PaM surroundings in coherence with our data and literature [61, 65]. We further validated some of these findings with immunohistochemistry (IHC), in FFPE and/or in PFA AD hippocampal sections. We confirmed the enrichment of STING in CoM, as well as C4d, a cofactor of C4B, and ErbB4, a co-receptor of ErbB2 [46] in PaM and PaM surroundings (Fig. 4C–E). Specifically, STING expression was elevated in AD hippocampal samples (8 AD, 7 CTL, FFPE), often associated with blood vessels and microglia-like cells in the white matter of the stratum oriens, CA3, and particularly the PL of CA1 (Fig. 4C). Often microglia aggregated around pyramidal neurons of the CA1 were STING immunoreactive. C4d staining was enriched around Aβ plaques in the hippocampus (CA3, CA1 and DG), subiculum, entorhinal and temporal cortex in a set of our samples (3 AD, 3 CTL, FFPE), validating previous observations [90] (Fig. 4D). Our 3D confocal analysis revealed the presence of some ErbB4-positive PaM encapsulated by GFAP-positive astrocytes in PFA-fixed hippocampal sections (Fig. 4E, Supplementary Fig. 5). These observations highlight distinct functional states and pathological microenvironments between PaM and CoM.
Fig. 4
Differentially expressed proteins between PaM and CoM and their direct microenvironment. A Cell-type enriched spatial protein profiling workflow. One slide with 5µm-thick FFPE hippocampal sections from three AD patients (cases #49, #50 and #52) was processed for SP using Nanostring GeoMx DSP technology. SP AOIs of PaM and CoM were selected based on Iba1-segmented areas. The remaining Iba1-negative area was used to assess the surrounding of PaM and CoM. B Volcano plots of DEPs in microglia PaM vs. CoM (left) and between their, respectively, surrounding (right). AOIs of Thresholds of Log2FC = 0.5, P value = 0.05 and FDR = 0.1 are indicated in the graphs. C–E Validation of proteins found enriched in PaM, CoM and their surroundings. C STING expression (DAB, brown) in microglia is upregulated in AD vs. CTL and found in various microglia population among them, microglia in CA1 PL seemingly aggregated in a CoM-like conformation around neurons (cases #14 and #36). D C4d (DAB, brown) is enriched in plaques vicinity in case #36. E 3D confocal images show erbB4 (magenta) expression in PaM (case #45). Scale bars: A 100 µm; C–E 30 µm
Digital spatial transcriptomic profiling reveals distinct and complex molecular and metabolic states of PaM vs. CoM and surrounding astrocytesTo unravel the molecular profiles and functional states of PaM and CoM and their respective immediate surrounding cells, we used the GeoMx DSP transcriptomic platform to quantify gene expression (18,677 RNA targets) in Iba1-positive PaM and CoM, and their respective surrounding GFAP-positive astrocytes (Fig. 5A). In addition, we also profiled some Rod, defined by their elongated morphology, and sometimes clustered in CoM-Rod (Supplementary Fig. 6A). We assessed a total of 38 AOIs across two consecutive sections from three AD patient samples (cases #49, #50 and #52): 10 PaM-Iba1, 10 PaM-GFAP, 7 CoM-Iba1, 7 CoM-GFAP, and 4 Rod-Iba1 AOIs. Under UV illumination, the indexing oligonucleotide tags were collected from the probe mix and counted individually from each Iba1-positive and, their respective, GFAP-positive AOI. Briefly, all AOIs passed sequencing quality control and targets with expression in less than 5% of the segments above the LOQ were removed. The target count matrix was scaled to the geometric mean of the nuclei and normalised by the upper quartile (Q3) normalisation. A final dataset of 15,518 genes was analysed.
Fig. 5
Spatial RNA profiling analysis unveil sets of differentially expressed genes between CoM and PAM and their surrounding astrocytes. A Cell-type enriched spatial transcriptome profiling workflow. Two slides, each with 5µm-thick FFPE hippocampal sections from three AD patients (cases #49, #50 and #52) that were cut at different depth levels of the bloc, were processed for ST using Nanostring GeoMx DSP technology. ST AOIs of PaM and CoM were selected based on Iba1-segmented areas and their surroundings based on GFAP-segmented areas. ST AOIs of Rod-shaped microglia were selected based on their bipolar morphology. B Venn diagrams representing overlap (total number and percentage) of top 500 genes for microglia AOIs of PaM, CoM and Rod, respectively top 500 genes for astrocyte AOIs of PaM and CoM. C Heatmaps of unsupervised clustering of DEGs between microglia AOIs (1,238 DEGs) and astrocyte AOIs (895 DEGs). D Volcano plots of DEGs in microglia AOIs of PaM vs. CoM. Thresholds of Log2FC = 0.5, P value = 0.05 and FDR = 0.1 are indicated in the graphs. E Heatmaps of top 30 DEGs in microglia AOIs PaM vs. CoM. Log2FC values are indicated in brackets for each transcript. F Volcano plots of DEGs in PaM vs. CoM astrocytes AOIs. G Heatmaps of top 30 DEGs between PaM and CoM astrocytes. H ORA GO of upregulated DEGs in PaM, CoM, PaM astrocytes and CoM astrocytes show distinct molecular pathways
Comparative analysis of the 500 most highly expressed genes in each microglial and astrocyte subgroup revealed overlaps and unique gene sets. We found an overlap of 197 genes (20.6% of total genes) between all the three groups of microglia, and 290 genes (40.8%) between the two groups of astrocytes (Fig. 5B). Approximately 20% of genes were exclusive to one specific group for microglia and 30% for astrocytes. Among the highly expressed genes in all microglial subgroups were allograft inflammatory factor 1 (AIF-1), CD74, or colony-stimulating factor-1 receptor (CSF-1R).
In contrast, astrocytic subgroups exhibited expression of astrocyte-specific markers such as GFAP, aldolase C fructose-bisphosphate (ALDOC), and ezrin (EZR). We then analysed the DEGs between the microglia or astrocyte subgroups using unpaired t test and multiple comparison adjustment (Benjamini–Hochberg with FDR of 0.1). We identified 1,238 DEGs in our microglia dataset and 895 DEGs in our astrocyte dataset. Unsupervised clustering resulted in a clear separation of all AOIs into morphologically predefined groups for the microglia and astrocyte datasets, as shown in the heatmaps (Fig. 5C). DEGs were displayed in volcano plots for all comparisons (Fig. 5D, Supplementary Fig. 6B-C for Rod) and the top 30 DEGs identified in heatmaps (most enriched and lowest p value) (Fig. 5E, G, Supplementary Fig. 6D-E). When comparing PaM to CoM transcripts, we found RAB27B, RHOQ, PCED1B, TMED8, HCAR1, CLVS1, GOLPH3, PTPRT, DCHS1, TNFAIP1, FOXO1, OXCT2, and HMCES among the most enriched and statistically significant genes for PaM; while in CoM they were C9orf131, TBXT, CASP2, AICDA, E2F6, ATP5F1D, MAP2K5, GIPC3, PPP1R9A, FBXL15, ACADS, ZMAT2, GSDME DUS3L, GDF1, and BLOC1S3. Rod-shaped microglia also showed numerous DEGs compared to both CoM and PAM. TAAR1, BCKDHA, CTNNAL1, EDIL3, RYR1, CALML4, RAX2, SLC20A2 and SERPINB6 were among the top 20 most DEGs in both comparisons (Supplementary Fig. 6B-C, Supplementary Excel 1).
We then performed an over-representation analysis (ORA) for PaM and CoM DEGs. The Gene Ontology (GO) pathways over-represented in PaM revealed phosphorylation and lipid degradation activities, antioxidant and glutathione transferase and tyrosine kinase activities. Consistent with this, the ORA based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) showed associations with sphingolipid signalling pathways, insulin resistance, ErbB and Wnt signalling pathways or necroptosis (Supplementary Excel 2) (Fig. 5H). The top five GO pathways for CoM were associated with ubiquitin-like protein transferase, purinergic nucleotide receptor, oxidoreductase cyclin-dependent kinase activities (Fig. 5H). ORA KEGG and Wikipathways (WP) suggested associations with transforming growth factor (TGF)-β and NF-κB signalling, among others (Supplementary Excel 2).
Next, we analysed DEGs in PaM astrocytes vs. CoM astrocytes. PaM astrocytes were associated with distinct signatures, and we found GOLGA8F, PCSK9, HOXD13, PIWIL1, PIK3C3, FAM71E2, FLG2, GBA2, SLC10A5, ACE, FBXO10, ARHGAP29, CES1, TMEM273, and GALNT7 among the most enriched genes (Fig. 5F, G, Supplementary Excel 1). We found SERPINA5, TUBB2A, FAM171A1, PROB1, GP1BA, TMEM37, FNDC4, JUNB, SCN5A, TSC22D4, GFPT1, BASP1, and UBE2W among the most enriched and statistically significant genes for CoM astrocytes. We performed ORA on PaM astrocytes and CoM astrocytes DEGs. ORA GO of DEGs PaM astrocytes showed enrichment for fibroblast growth factor receptor signalling, response to TGF-β, glucose transmembrane transport, positive regulation of apoptotic process and amyloid beta clearance (Supplementary Excel 2). ORA KEGG and WP indicated activities associated with PI3K-Akt signalling pathway, cholesterol metabolism or cell adhesion molecules, fatty acid and lipoprotein transport or complement system. ORA GO highlighted the association of DEGs CoM astrocytes with metabolite and energy generation (OXPHOS), cytokine mediated pathways but also with adaptive immune response, JAK-STAT signalling and cellular response to misfolded proteins (Fig. 5H, Supplementary Excel 2). Our DSP analysis revealed specific cellular and molecular identities of PaM and CoM, and of their respective surrounding astrocytes.
Markers of the molecular signatures of PaM, CoM and surrounding astrocytes in the hippocampus of AD patientsTo validate the molecular signatures identified in PaM and PaM astrocytes by our DSP analysis, we performed IHC on selected proteins that embodied an enriched pathway in FFPE or PFA samples. First, we examined the complement component 1q (C1q) (n = 5 AD, 5 CTL) and complement component 3 (C3) (n = 3 AD, 2 CTL) association with PaM and the PaM environment or PaM astrocytes. In AD, C1q was strongly distributed around plaques, and on some hippocampal neuronal cell bodies and neurites in the CA3-CA1 subfields, but also in the DG (Fig. 6A) and entorhinal cortex (Supplementary Fig. 6A), and occasionally diffuse in areas reminiscent of astrocyte domains. C1q-positive hippocampal neurons were also found in CTL (Supplementary Fig. 7A). Similarly, C3 staining exhibited lower overall expression, primarily labelling the core and corona of Aβ plaques in AD (Fig. 6B, Supplementary Fig. 7B). Using high-resolution 3D confocal imaging, we observed C3 expressed within GFAP-positive PaM astrocytes territories, in cell bodies but also along processes (Fig. 6C). We then analysed the distribution of Proprotein convertase subtilisin/kexin type 9 (PCSK9) (n = 3 AD, 2 CTL) and Integrin subunit alpha 6 (ITGA6, also known as CD49f [6]) (n = 5 AD, 5 CTL), associated with the PaM astrocytes and PI3K-Akt pathways in our previous analysis. In FFPE sections, we observed a PCSK9 expression in neurons both in CTL and AD (Supplementary Fig. 7); however, in AD samples, PCSK9 staining was also associated with plaque microenvironments and with cells resembling PaM astrocytes in hippocampus and entorhinal cortex (Fig. 6D, E, Supplementary Fig. 7). ITGA6 was predominantly distributed along blood vessels (BV) in CTL, while in AD, expression around BV was weaker and occasionally observed near plaques, apparently associated with PaM astrocytes. We confirmed a low expression of ITGA6 in the cell bodies and processes of PaM astrocytes with confocal microscopy (Fig. 6F). We then investigated the association of the PaM microenvironment with necroptosis identified through pathway analysis. We found, in bright-field and 3D confocal images, a clear enrichment of phosphorylated mixed lineage kinase domain-like protein (pMLKL), part of the necrosome complex, in PaM vicinity but no staining in CoM (Fig. 6G). PMLKL was frequently concentrated in vacuole-like accumulations within the cytoplasm of numerous pyramidal neurons in AD samples and, to a lesser extent, in CTL as previously described [42].
Fig. 6
Validation of functional markers for PaM, PaM astrocytes and CoM by IHC on human post-mortem hippocampal samples. A C1q (DAB, brown) is expressed in the vicinity of plaques, in cells resembling microglia in CA1 AD (case #30). Some pyramidal neurons were also stained by C1q in AD. B, C C3 (DAB, brown) is also expressed close to plaques (cases #38 and #40). C In confocal microscopy, C3 (magenta) punctuated distribution is found within PaM astrocyte territories (GFAP, green). D PCSK9 (DAB, brown) which is mainly expressed by neurons is enriched in the plaque vicinity in AD (case #44). E, F ITGA6 is associated with blood vessels and plaque microenvironments (cases #14, #33 and #35). G IHC fluorescent high-resolution confocal 3D acquisitions of a post-mortem AD hippocampus showed an enrichment of the necroptosis marker pMLKL (green) in the vicinity of plaques (full arrows), while CoM (empty arrows) were not engulfing pMLKL + structures (left panel 3D, right panel z focal plan) (case #29). H, I NF-κB p65 (orange) nuclear translocation was found at high level in CoM (empty arrows) (H), in PaM (full arrows), and PaM astrocytes (I) in AD samples (case #25). J Confocal 3D images show that CoM (Iba1, magenta) express some SMURF2 (green) proteins (cases #52, # 35). K Chromogenic staining show the distribution of ACADS (DAB, brown) in the CTL and AD hippocampi (cases #15 and #37). L Confocal 3D images show that ACADS (green) expression is found in CoM (Iba1, magenta) (cases # 35). Scale bars A–F, H, I, K, L 30 µm; G 40 µm (low mag), 10 µm (high mag); J 30 µm (low mag), 15 µm (high mag)
DSP analysis suggested that CoM was involved in different biological processes than PaM. To further elucidate these processes, we performed co-immunostaining against Iba1, GFAP and NF-κB -p65 in PFA sections revealing strong expression and translocation of NF-κB -p65 into CoM nuclei by confocal microscopy (Fig. 6H). The staining was not exclusive to CoM, but also detected in the surrounding microenvironment, without being predominantly expressed in astrocytes. In contrast, PaM and PaM astrocytes displayed heterogeneous NF-κB -p65 staining patterns (Fig. 6I). ORA analysis revealed strong ubiquitin ligase activity in CoM. Confocal microscopy analysis of E3 ubiquitin-protein ligase SMURF2 distribution in AD PFA-fixed samples revealed expression in CoM, although not significantly enriched compared to the surrounding microenvironment (Fig. 6J). Acyl-CoA dehydrogenase (ACADS) is associated with fatty acid oxidation and high metabolic activity. In CTL samples, ACADS was expressed in astrocytic cells in the temporal cortex, CA4 hippocampus and CA3-CA1 PL. In AD, astrocytes retained ACADS expression, but so did some microglia surrounding pyramidal neurons. 3D confocal analysis confirmed the co-distribution of ACADS in the CoM (Fig. 6K, L).
Macrophage and T cell association with PaM and CoMTo determine whether infiltrating immune cells contribute to the PaM or CoM molecular markers and composition, we next examined the distribution of T cells and CNS-associated macrophages in CTL and AD hippocampal samples. Indeed CoM exhibited a cellular organisation reminiscent of nodules observed in various brain infections, often containing T cells [80, 95]. First, we quantified the number of CD8 + T cells, which have previously been detected near Aβ plaques in AD brain samples, in CTL and AD hippocampi (4CTL, 6 AD). CD8 + T cells were mainly found in BV and less frequently in hippocampal parenchyma (Fig. 7A). CD8 + T cells were slightly increased in AD without reaching significance (Fig. 7B). To further characterise the properties and distribution of CD8 + T cells, we used multiplex chromogenic IHC to stain CD3 in combination with CD8, together with 4G8 for Aβ plaques and Iba1 for microglia in some CTL and AD hippocampal samples (CTL n = 3; AD n = 6). With this approach, we detected only single positive cells, mainly CD8 + and sometimes CD3 + (Fig. 7C, D). In CTL hippocampus, CD8 + cells were strongly localised in the BV, with occasional presence in the parenchyma, close to microglia. In AD, we again observed a slight increase in parenchymal CD8 + cells, sometimes infiltrating in hotspots, but with very little to no association with PaM, Aβ plaques or CoM (Fig. 7D). Similar observations were made in the entorhinal and temporal cortices of AD samples (Supplementary Fig. 8A).
Fig. 7
Comments (0)