
Remember me





LncRNA transcripts of more than 200 nucleotides that are not translated into proteins. It is widely acknowledged that Chemoresistance and cancer biology are both affected by long non-coding RNAs (lncRNAs). They also play key roles in the pathogenesis and progression of LC. Several mechanisms are involved in these effects, such as epigenetic regulation, which affects cell proliferation and differentiation [10, 11]. Abnormally expressed lncRNAs are recognized as significant contributors to the development of LC and they offer promise as diagnostic, therapeutic, and prognostic biomarkers. Over the past few years, researchers have identified and extensively studied many lncRNAs in LC [12].
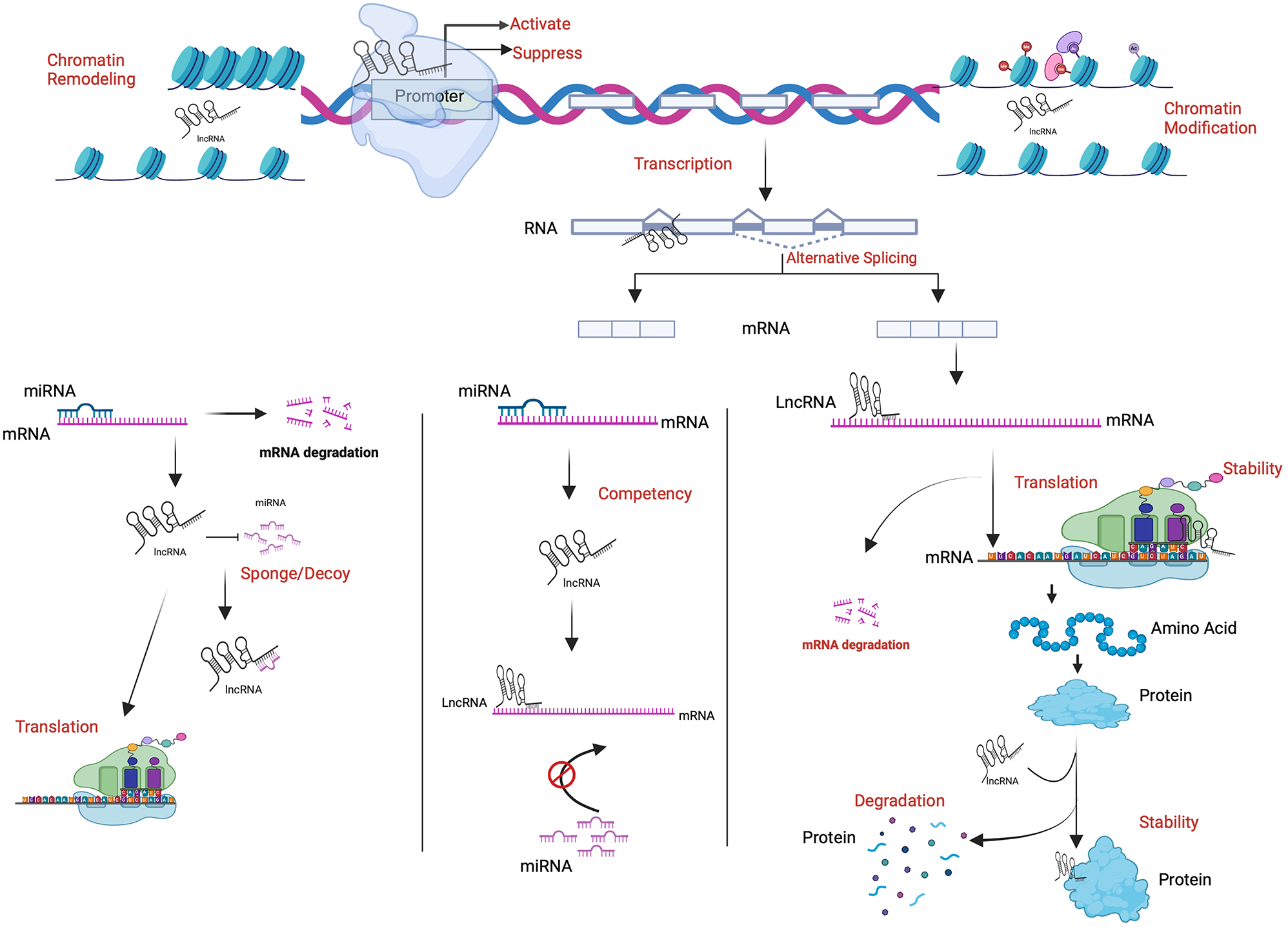
Mechanisms of lncRNA-mediated chemoresistanceDespite the development of therapeutic strategies for LC, resistance to therapy is inevitable. Increasingly, LC treatments face the challenge of chemoresistance, and genetic changes appear contribute significantly to this process. Several mechanisms have been identified by which lncRNAs regulate chemoresistance in cancer cells. Some broad categories of mechanisms are discussed in the following sections (Fig. 1).
Fig. 1
Schematic illustration of the mechanisms by which lncRNAs affect chemoresistance. This figure was designed with BioRender.com
Direct impact of lncRNA on gene expressionAn important mechanism by which lncRNAs influence drug resistance is through their ability to modulate the expression of related genes. Based on their subcellular localization, lncRNAs directly affect gene expression. lncRNAs engage in chromatin modification by binding to chromatin regulatory proteins, directly influencing gene regulation, altering the splicing and stability of mRNAs, and indirectly participating in transcriptional and post-transcriptional regulatory mechanisms by interacting with other RNAs and proteins [13,14,15]. The activities of lncRNAs, such as metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), maternally expressed 3 (MEG3), and H19, play important roles in the regulation of the cell cycle, particularly through their effects on key proteins, such as p21 and p53 [16,17,18]. Alternatively, lncRNAs, such as H19, pituitary tumor-transforming (PINT), lincRNA-p21, and PPARα-binding non-coding RNA in diabetes (PANDA), are upregulated by p53, showing the complex interaction between lncRNAs and major regulatory pathways [18,19,20,21,22].
HOX transcript antisense RNA (HOTAIR) illustrates how lncRNAs function as both enhancers and inhibitors of gene expression, demonstrating the dual roles of lncRNAs in gene regulation. By interacting with chromatin-modifying complexes, HOTAIR alters the chromatin structure, which in turn affects gene transcription. Specifically, HOTAIR recruits polycomb repressive complex 2 (PRC2) and LSD1/CoREST/REST complexes to target genomic locations [23]. By modifying histones, including methylation and demethylation, these complexes facilitate chromatin condensation and suppress the expression of certain target genes, while activating others associated with resistance mechanisms.
In addition to modulating chromatin states, HOTAIR also influences the binding affinity of transcription factors to DNA, which controls the transcription of several genes essential for drug resistance, including those that inhibit apoptosis and drug efflux and support cell survival [24]. The regulatory capabilities of HOTAIR in cancer, particularly those related to cellular survival and drug metabolism, contribute to a resistant phenotype, while its expression in breast and liver cancers is affects clinical outcomes and induces resistance to chemotherapy [25]. HOTAIR plays an important role in the orchestration of complex epigenetic programs that may serve as targets for therapeutic approaches aimed at overcoming cancer-related resistance.
lncRNAs modulate the activity of RNA-binding proteinsAnother mechanism by which lncRNAs regulate chemoresistance is by modulating the activity of RNA-binding proteins (RBPs), which are essential for mRNA stability, translation, and splicing, thus influencing the expression of proteins related to drug resistance. In cellular systems, lncRNAs and RBPs play key roles in regulating gene expression. The interaction between these two genes is key to the post-transcriptional regulation of mRNA and directly affects the synthesis of cancer-relevant proteins by regulating mRNA stability [26]. Researchers have demonstrated that RBPs modulate mRNA stability via post-transcriptional mechanisms, which are crucial for the initiation and progression of malignant tumors and their resistance to treatment [27, 28]. It is important to note that, although RBPs typically bind to mRNAs, they can also interact with lncRNAs. Both lncRNAs and RBPs exhibit abnormal expression patterns in tumors, which may alter tumor behavior and responses to treatment [29, 30]. Recent studies have shown that lncRNAs regulate mRNA stability by interacting with RBPs, which significantly contributes to cancer progression and drug resistance [31,32,33].
lncRNAs are significant regulators of post-translational modifications of RBPs. PLK1 is phosphorylated and activated by anaplastic lymphoma kinase (ALK) 5’ UTR pseudogene –associated lncRNA (APAL), which is vital for the survival of NSCLC cells. One study highlighted the crucial role of lncRNAs in cancer cell survival and identified APAL as a potential therapeutic target for a variety of cancers. They found that APAL knockdown caused mitotic catastrophe and substantial apoptosis in human lungs. In contrast, APAL overexpression accelerated cell cycle progression, enhanced proliferation, and inhibited the induction of apoptosis by chemotherapy. Based on the results of mechanistic studies, it appears that APAL facilitates the interaction between PLK1 and Aurora A, which in turn enhances the ability of Aurora A to phosphorylate PLK1. In vivo experiments have shown that targeting APAL inhibits LC xenografts [34].
lncRNAs can cause the intracellular localization of RBPs. For example, the lncRNA AC020978 interacts with PKM2 and promotes its translocation from the nucleus to the cytoplasm, resulting in enhanced proliferation and glycolytic metabolism in NSCLC [35]. According to the findings of the that study, AC020978 was significantly upregulated in NSCLC and its upregulation was associated with advanced TNM stage and poor clinical outcomes, indicating that it can serve as an independent prognostic indicator. In NSCLC, AC020978 confers an aggressive phenotype and poor prognosis. Thus, it may be beneficial to target AC020978 in the treatment of patients with NSCLC [35].
lncRNAs interact with microRNAsIn addition to other mechanisms underlying the regulation of chemoresistance by lncRNAs, lncRNAs can function as molecular sponges for microRNAs (miRNAs), thus inhibiting their activity. The ability of lncRNAs to sequester miRNAs prevents them from repressing their target mRNAs, including genes associated with survival and resistance to chemotherapy. As competing endogenous RNAs (ceRNAs), lncRNAs can indirectly modulate mRNA expression via a sponge mechanism [36]. It has been shown that lncRNA-mediated sponge interactions and their protein-coding targets can be involved in various malignancies, including gastric cancer [37], glioblastoma multiforme [38], pancreatic cancer [39], ovarian cancer [40], and breast cancer [41].
As a prominent miRNA sponge, the parentally expressed lncRNA H19 indirectly regulates the expression of downstream target genes, thus facilitating cancer progression in a variety of tumor types. Within a single cancer type, H19 exhibits versatility by sponging various miRNAs to mediate different regulatory outcomes [39]. Due to its highly conserved secondary structure, H19 can bind to miRNAs and proteins, allowing it to act as a ceRNA. Furthermore, H19 can recruit and bind to the enhancer of zeste homolog 2 (EZH2) or function through its derivative miRNA-775 to inhibit the expression of target genes [42]. Recent studies have recently highlighted the role of H19 as a ceRNA that indirectly regulates downstream mRNAs that play critical roles in promoting or inhibiting tumorigenesis. According to one study [43], H19 regulates cisplatin resistance in human lung adenocarcinoma cells.
Based on a recent study, lncRNA small nucleolar RNA host gene 14 (SNHG14) is upregulated in NSCLC cells and is responsible for resistance to chemotherapy treatments involving cisplatin. Upon silencing SNHG14, NSCLC cells were found to be significantly more sensitive to cisplatin. By decreasing miRNA-133a expression levels, SNHG14 promotes the expression of the homeobox protein Hox-B13 (HOXB13) [44]. NSCLC cells are sensitized to cisplatin when doublecortin-like kinase 1 (DCLK1) is downregulated by miRNA-330-5p. SNHG1 downregulates miRNA-330-5p expression and upregulates DCLK1 to exert tumor-suppressive activity Accordingly, silencing SNHG1 reverses the resistance of NSCLC cells to cisplatin. There is also evidence that the lncRNA X-inactive specific transcript (XIST) interacts with miRNA-101-3p in LC [45]. Overexpression of XIST in lung tumors promotes cisplatin resistance by glucose uptake, acidification rates, and lactate production, and by inducing glycolysis enhances the growth of lung tumors. Through its role as a ceRNA, XIST inhibits the expression of miRNA-101-3p and contributes to cisplatin resistance [45]. Restoration of miRNA-101-3p expression can abrogate the tumor-promoting function of XIST in LC and increase the sensitivity of LC to cisplatin. In addition to miRNA-101-3p, XIST also regulates the response of NSCLC cells to chemotherapy by regulating miRNA-520. Through its ability to sponge miRNA-520, XIST increase cisplatin resistance and inhibit apoptosis by modulating Bcl-2-associated X (BAX) expression and the p53 pathway [46].
lncRNAs regulate biological processesAdditionally, lncRNAs have been increasingly observed as pivotal regulators of various biological processes, including cell survival, apoptosis, and drug metabolism. A growing body of evidence supports the contribution of lncRNAs, such as XIST, to the progression and metastasis of cancer. XIST is dysregulated in various tumor types, including NSCLC. According to in vitro studies, knockdown of XIST inhibits the proliferation of NSCLC cells and enhances their sensitivity to cisplatin (DDP) through apoptotic and pyroptotic pathways [47]. The oncogenic properties of XIST and its ability to facilitate DDP resistance have been attributed largely to its interaction with the TGF-β effector protein SMAD2. This interaction prevents SMAD2 from translocating to the nucleus, thereby inhibiting p53 and NLRP3, two key transcription factors that regulate the apoptosis and pyroptosis processes, respectively. Based on experiments using DDP-resistant NSCLC cell lines and mouse xenograft models, it can be concluded that XIST is oncogenic and inhibits programmed cell death, thereby increasing DDP chemoresistance [47].
Growth arrest-specific transcript 5 (GAS5) is an lncRNA that promotes apoptosis of cancer cells. This primarily affects the sensitivity of the LC to chemotherapy. GAS5 promotes cisplatin sensitivity in LC by inhibiting autophagy [48]. It also promotes gefitinib-induced LC cell death by inhibiting insulin-like growth factor 1 receptor (IGF-1R) [49]. Furthermore, maternally expressed 3 (MEG3) increases the sensitivity of LC cells to cisplatin by increasing p53 and β-catenin levels and cell survival [50]. In LC, experiments using nuclear enhanced abundant transcript 1 (NEAT1) revealed that the interaction between NEAT1 and copper transporter 1 (CTR1) facilitated the internalization of cDDP (platinum-based chemotherapies, such as cisplatin) in tumor cells, thereby increasing cisplatin sensitivity [51].
lncRNAs modulate signaling pathwaysOne way that lncRNAs exert their regulatory effects is by modulating critical signaling pathways that contribute to the development of chemoresistance. The relationship between lncRNAs and cancer regulatory pathways has been studied previously, but few comprehensive studies have been conducted [52]. Several malignant diseases, such as breast cancer [53], prostate cancer [54], gastric cancer [55], and pancreatic cancer [56] are constitutively activated via the PI3K/Akt/FOXO and NF-B pathways [56].
For example, the lncRNA Linc00152 has been shown to promote gastric cancer growth by activating the EGFR-dependent PI3K/Akt pathway [57]. Similarly, the lncRNA BC087858 enhances LC invasion and drug resistance to epidermal growth factor receptor (EGFR) inhibitors by activating the PI3K/Akt pathway [58]. Through the same signaling cascade, the lncRNA MALAT1 has been implicated in cholangiocarcinoma progression [59].
A positive feedback loop between the lncRNA plasmacytoma variant translocation 1 (PVT1) and Wnt/β-catenin signaling has been documented in the context of gemcitabine resistance in pancreatic cancer. In addition to providing insights into the mechanisms by which lncRNAs regulate autophagy, this study demonstrated how lncRNAs interact with autophagy-related proteins. Such cooperation between lncRNAs, autophagy, and Wnt/β-catenin signaling pathways may mitigate the effects of chemotherapeutic stress [60].
During the past few years, researchers have begun to examine the effects of natural compounds on lncRNAs and how they interact with NF-κB signaling pathways. In addition, silencing of the lncRNA nuclear factor I/A-like protein (NKILA) negates the anticancer effects of baicalein, indicating that NKILA plays a critical role in mediating these effects. As well, the NF-κB nuclear translocation inhibitor JSH-23 was found to diminish the effects of NKILA, thereby establishing a connection between the actions of baicalein, lncRNA NKILA, and NF-κB signaling pathways [61].
There is evidence that the downregulation of HOTAIR promotes the sensitivity of cells to anticancer drugs, resulting in the suppression of cell viability, arrest of the cell cycle, and the prevention of tumor development [62]. There is also evidence that the lncRNA AC006050.3-003 plays a significant role in the development of chemoresistance [63]. In addition, AK126698 has been found to induce cisplatin resistance in LC cells by targeting the Wnt pathway [64]. Urothelial cancer-associated 1 (UCA1)-induced cancer cells acquire resistance to epidermal growth factor receptor tyrosine kinase inhibitors [65].
lncRNAs are currently under investigation for their involvement in chemoresistance in LC. Several lncRNA targets have demonstrated the potential to address the challenge of chemoresistance (Table 1).
Table 1 lncRNAs targets with promise to overcome the obstacle of chemoresistanceSeveral mechanisms appear to participate in the promotion of chemoresistance in LC, including the regulation of gene expression, activation of signaling pathways, and ceRNAs. Targeting these specific lncRNAs may improve treatment outcomes in patients with LC by overcoming drug resistance. Understanding the roles of lncRNAs in chemoresistance has advanced significantly; however, several challenges remain.
Exosomes: mediators of lncRNA-driven chemoresistance in lung cancerCells secrete nano-sized vesicles called exosomes, which play important roles in intercellular communication. Different molecules are transported by exosomes, including proteins; lipids; and nucleic acids, such as lncRNAs.
lncRNAs are transferred between cells by exosomes and they play an essential role in the development of chemoresistance in LC. Various cellular processes, including proliferation, differentiation, and survival, are regulated by lncRNAs. Moreover, lncRNAs contribute to resistance to chemotherapy and targeted therapies [66,67,68]. To promote chemoresistance, exosomes act as messengers that transfer lncRNAs from resistant tumor cells to sensitive cells.
Exosomal lncRNAs are involved in mediating drug resistance in LC. An example of an exosomal lncRNA is RP11–838N2.4, which can be packed into exosomes to decrease the sensitivity of cells to chemotherapy drugs [67]. Recent studies have identified two lncRNAs, maternally expressed gene 3 (MEG3) and ETS1-associated transcript 1 (MLETA1), high-metastasizing LC cells and their secreted exosomes are upregulated in these pathways. Moreover, lncRNA MLETA1 plays a role in promoting LC metastasis and cell motility by regulating the expression of EGFR and insulin-like growth factor 1 receptor (IGF1R) and sponging miR-186-5p and miR-497-5p.1 [66].
UFC1, also known as urothelial cancer-associated 1 (UCA1), is an exosomal lncRNA that inhibits apoptosis and cell cycle arrest and induces the proliferation and metastasis of lung tumor cells. EZH2 accumulates at the promoter of phosphatase and tensin homolog (PTEN) as a result of its of interaction with EZH2. A significant reduction in PTEN expression leads to NSCLC progression [69]. Moreover, investigated the relationship between exosomal lncRNA H19 and erlotinib resistance in LC. H19 can be loaded into exosomes and transferred to LC, which may reduce sensitivity to erlotinib. To achieve this goal, exosomal lncRNA H19 inhibits miRNA-615-3p expression to enhance autophagy-related 7 (ATG7) expression in LC cells, thereby causing the cells to become resistant to erlotinib [70].
As research emphasizes the role that interactions between tumors and stromal cells play in immune evasion and cancer metastasis, attention is being paid to the tumor microenvironment [71]. Among the main components of the tumor microenvironment are cancer-associated fibroblasts (CAFs), which are involved in the process of promoting tumor development by secreting exosomes, extracellular membrane vesicles, DNA, and various forms of RNA, which serve as messengers between cells [72]. A number of studies have demonstrated that CAF-derived exosomal lncRNAs are involved in the progression of cancer and immune evasion [73,74,75,76].
MEG3 released from CAF-derived exosomes confers DDP resistance via regulation of the miR-15a-5p/CCNE1 axis in SCLC. The current study may provide a new potential therapeutic strategy for improving the clinical benefits of DDP. The combination of etoposide/topotecan plus DDP has been shown to be effective as the first-line chemotherapy for patients with SCLC [77]. Several additional exosomal lncRNAs involved in the regulation of LC progression are shown in Table 2.
Table 2 The exosomal lncRNAs in regulating progression of lung cancer cellsTherapeutics based on targeting lncRNAs: a new approach in drug discoveryTargeting lncRNAs with drugs is a promising research topic. An lncRNA-targeted therapeutic approach aims to enhance therapeutic effects by modulating lncRNA expression or function. lncRNAs are involved in cancer progression and tumorigenesis at multiple stages and are widely expressed in LC [78]. These molecules play cretical roles in the development and regulation of a wide variety of molecular pathways associated with gene expression. Molecular-targeted therapies and chemotherapy have been reported to be associated with the dysregulation of lncRNAs. Since they are specific and sensitive to chemotherapeutic drugs, lncRNAs may serve as new therapeutic targets for NSCLC and may prove to be effective at curing it [79]. The methods employed to address chemoresistance by degrading, inhibiting, or modifying lncRNAs are presented in Fig. 2.
Fig. 2
Examples of advances in targeting lncRNAs and the potential therapeutic benefits of this approach. Antisense oligonucleotides (ASOs) and small interfering RNAs (siRNAs) are capable of degrading long non-coding RNA (lncRNA) molecules in different ways. (a) An ASO is a synthetic nucleic acid strand that is designed to bind specifically to the complementary bases of an lncRNA. Binding of the ASO-RNA complex to a lncRNA facilitates degradation by RNase H, as RNA-DNA hybrids are cleaved by RNase H. (b) siRNAs are usually short double-stranded RNA molecules, with one strand matching the sequence of the target lncRNA. After entering the cell, the siRNA is incorporated into an RNA-induced silencing complex (RISC). The lncRNA is degraded once it has been bound by the Argonaute protein component of the RISC. (c) Steric inhibitors, such as small molecules, morpholinos, or uniformly modified ASOs, can be used to inhibit lncRNA-protein interactions. ASOs bind to nascent lncRNA transcripts. Consequently, mature lncRNA levels can be reduced, and their activity can be effectively inhibited. A morpholino is a synthetic molecule that binds to a lncRNA and prevents other molecules from accessing the sequence. This method is stable and resistant to nucleases, allowing it to effectively inhibit lncRNA expression without degrading the target lncRNA. (d) CRISPR/Cas9 technology provides an efficient and flexible means of modulating lncRNA expression levels directly by targeting genomic DNA. Guide RNAs (gRNAs) are designed to match sequences upstream or within genes that encode the lncRNAs. The Cas9 endonuclease is directed by the gRNA to the specific location in the genome where double-strand breaks are generated. In the event that these breaks are introduced, insertions or deletions (indels) may be generated during the DNA repair process, typically as a result of non-homologous end joining. BioRender.com
With recent advancements in genome editing technologies, such as clustered regularly interspaced short palindromic repeats (CRISPR)-associated protein 9 (CRISPR-Cas9) technology, it has become possible to silence the transcription of lncRNA genes through CRISPR interference [156, 157]. Using this technique, a transcriptional-repressor-fused dead-Cas9 protein is directed to a specific gene promoter via guide RNAs to silence the gene [158]. CRISPR/Cas9 has been used to target nuclear-enriched abundant transcript 1 (NLUCAT1), which is constitutively upregulated in lung adenocarcinomas under oxidative stress and hypoxic conditions. This results in a decrease in cell proliferation and invasion, as well as an increase in sensitivity to cisplatin-induced apoptosis [80]. Induced pluripotent stem cells and cancer cells are represented by seven human cell lines, were used to selectively deactivate lncRNAs via CRISPR interference. Approximately 500 lncRNAs were found to be critical to the proliferation of cancer cells. Most of these lncRNAs identified as essential for only one cell type, underscoring the specificity of their functions [159]. Knockdown of six different lncRNAs has been achieved using dCas9-KRAB, with > 80% efficiency observed for five lncRNAs, according to Gilbert et al. [81].
Loss of function may also be accomplished by steric inhibition of RNA-protein interactions or by inhibiting the formation of secondary structures. An RNA-binding small molecule or an antisense oligonucleotide (ASO) can be applied in this situation [82].
An effective method for targeting and degrading lncRNA is to use small interfering RNAs (siRNAs) or ASOs. Pathogenic RNAs can be knocked down via post-transcriptional RNA degradation. The cleavage pathway can be triggered by Dicer- and argonaute-dependent siRNAs. In addition, chemically modified ASOs can be used to target RNAse-H-dependent mechanisms to degrade the RNA of interest [82]. Targeted knockdown of MALAT1 and ANRIL in LC cells reduces cell proliferation and increases apoptosis [
Comments (0)