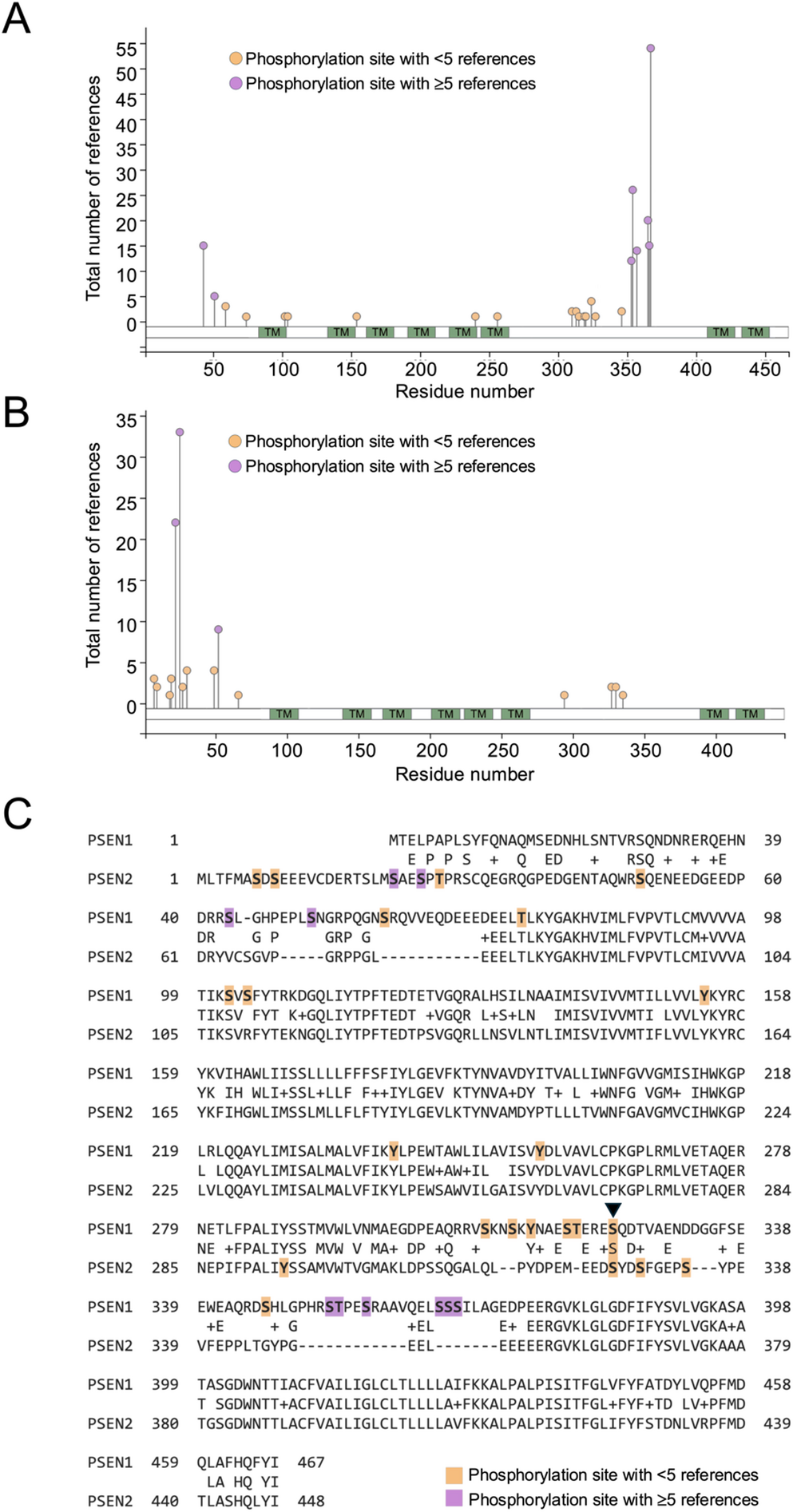
The resulting data for canonical serine, threonine or tyrosine phosphorylation is presented as lollipop plots in Fig. 1A and B, with detailed information for each modification available in supplemental Tables 1 and 2 and via the Phosphosite portal. Strikingly, only one of the reported phosphorylation sites (residue S324 in Presenilin 1 and residue S327 in Presenilin 2) is conserved across both proteins, with all other sites being unique to one or other of the proteins (Fig. 1C). This divergence is consistent with the primary sequence organization of the proteins.
To gain an insight as to the orientation of these residues within the Presenilins, the amino acids modified by phosphorylation were identified in simplified ribbon diagrams for Presenilin 1 and Presenilin 2 (Fig. 2). Although atomic resolution cryogenic electron microscopy (cryoEM) structures have been derived for gamma secretase containing both Presenilin 1 and Presenilin 2, it is of note that the residues catalogued in this study as being phosphorylated all sit in portions of their primary sequence that are not visible in these structures (PDB 5A63 and PDB 7Y5Z). This is likely due to their residing in flexible portions of the gamma secretase complex that are not amenable to visualisation by cryoEM.
The pattern of phosphorylation observed across Presenilin 1 and 2 is striking, and has some important implications. Most notably, and surprisingly, there is a clear divergence between the two Presenilins with regard to the location of phosphorylation sites. As noted above, Presenilin 1 and 2 are paralogs, sharing 63% identity and 7% similarity, and have a shared and redundant function - at least at a biochemical level - in the gamma secretase complex. This divergence is consistent with the primary sequence organization of the proteins. While Presenilin 1 and 2 are highly conserved within their transmembrane domains, many of the non-conserved phosphorylation sites map to cytosolic regions, specifically the N-terminal region and the large cytosolic loop. These regions exhibit reduced sequence conservation between Presenilin 1 and 2, likely contributing to the largely non-overlapping phosphorylation landscape by providing distinct kinase recognition motifs and regulatory interfaces in each paralog.
The human genetics of the PSEN genes indicates a closely aligned pathological role, with mutations in either gene resulting in a common phenotype. The observation that phosphoregulation of these proteins differs, however, suggests distinct roles within cells and tissues for each of the Presenilins, and implies that there are unique regulatory events governing these proteins. This opens up the possibility of specifically modulating the Presenilins individually via targeted inhibition and activation of upstream kinases – potentially indirectly modifying APP processing, and amyloid beta homeostasis through altered localisation, trafficking, or turnover of gamma secretase complexes, rather than direct changes in catalytic specificity.
Beyond the divergence in phosphorylation patterns, several studies have highlighted functional consequences of presenilin phosphorylation. Walter and co-workers reported that phosphorylation of Presenilin 2 near caspase cleavage sites reduces caspase-mediated proteolysis, delays apoptotic progression and protects Presenilin 2 from cleavage, whereas phospho-deficient Presenilin 2 is more readily cleaved (Walter et al., 1999). Similarly, Presenilin 1 phosphorylation has been shown to be critical for microglial phagolysosomal competence, including lysosomal acidification and efficient degradation of amyloid beta oligomers, effects that appear at least partly independent of canonical gamma secretase activity (Ledo et al., 2021). Mechanistic studies support these functional roles: for example, Presenilin 2 contains a phosphorylation-dependent AP-1 adaptor binding motif that directs Presenilin 2/gamma-secretase to late endososomes and lysosomes, a motif absent in Presenilin 1, explaining the preferential contribution of Presenilin 2 to APP processing in endolysosomal compartments (Sannerud et al., 2016). The observed sequence divergence in cytosolic loops, and the functional characterisation noted above, indicate that phosphorylation has been tailored to confer paralog-specific functional specialisation, regulating protein stability, trafficking stress responses and cell-type specific functions beyond classical gamma secretase cleavage activity.
There are a number of caveats to interpreting the phosphorylation events catalogued in this study. The underlying data represented on the Phosphosite portal includes both high-throughput unbiased analyses and low-throughput targeted analysis, however it is of note that only a minority of the phosphorylation sites across Presenilin 1 and 2 have been reported by more than five independent analyses. With decreased numbers of replication studies comes a decreased confidence in the physiological relevance of the phosphorylation event, a qualification that is relevant to the S324/S327 conserved phosphosites where there are 4 and 2 supporting references respectively. Several factors likely contribute to this limited overlap and validation: historical research bias, in which familial AD mutations in PSEN genes attracted the majority of attention toward studying mutational effects rather than post-translational regulation; technical challenges, since presenilins are low abundance, multipass membrane proteins that are difficult to solubilize and study by mass spectrometry; and context dependence, where phosphorylation events may be transient, cell-type-specific, activity-dependent, or stress-induced, limiting reproducibility across studies. The limited number of reports does not imply that these modifications are biologically irrelevant, as many phosphosites may exert context dependent effects on protein localisation, stability, or function. Consistent with this, Matz and co-workers identified 11 novel Presenilin 1 phosphosites but observed minimal effects on gamma secretase activity and abeta production under the tested conditions, suggesting that many of these phosphosites are permissive rather than universally instructive (Matz et al., 2015).
In addition, although Phosphosite compiles data on a wide range of post-translational modifications, encompassing acetylation, ubiquitylation and multiple classes of phosphorylation, this study has focused on reported canonical serine, threonine, and tyrosine phosphorylation events. This has the benefit of a wider evidence base for phosphorylation events, but has the drawback of missing the wider landscape of posttranslational modification as a means of regulating Presenilin function. A broader examination of posttranslational modification is beyond the scope of this current study, but is certainly merited in the future (especially in the light of rapidly increasing sensitivity and volume of proteomic analysis).
Finally, only a limited number of the phosphorylation events covered by this study have been characterised and validated in detail, with even fewer data available on the functional consequences of posttranslational regulation. Defining and understanding the signal transduction pathways that govern the phosphorylation of the Presenilins, and gaining a deeper comprehension of how these modifications alter Presenilin function, localization, stability, and interaction networks, should be prioritised in future investigations.


Comments (0)