Animals
In this study, wild-type C57BL/6J male mice (12–16 weeks old; CLEA Japan, Shizuoka, Japan). Animals were housed in rich-environment cages with extra bedding, a tunnel and a running wheel. Experiments were conducted in strict adherence to the approved guidelines by the Okinawa Institute of Science and Technology (OIST) and the Institutional Animal Care and Use Committee (IACUC), within a facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC International). 4 mice were used for anterograde labeling of SC neurons, 3 mice were used for retrograde labeling of IO afferents, 3 mice were used to label SC axons and sparsely label IO neurons, and another 3 mice were used for anterograde labeling of SC neurons combined with immunofluorescece. 2 mice were used for in-vivo calcium imaging experiment.
Surgical procedures for viral expression
Mice were anesthetized with 5 % isofluorane (delivered by means of the SomnoSuite vaporizer system, Kent Scientific, CT, USA) and placed in the stereotaxic frame (KOPF instruments, CA, USA) equipped with a heat pad (38\(^\)C; TMP-5b, Supertech Instruments). During surgery, the animal was kept under anesthesia with 1.5–2.5% isofluorane delivered through a nose cone. Precise alignment of the head and body is crucial for accurate targeting of deep structures such as the IO (Dorgans et al. 2022; Guo et al. 2021; Guo and Uusisaari 2025). The scalp was shaved and local anesthesia was administered using Xylocaine gel (Xylogel, gel 2%, Aspen, Japan), and the skin was disinfected and incised with a scissor to expose the skull. Subsequently, small craniotomies (approximately 1 mm in diameter) were opened using a handheld drill (Surgic XT Plus drill, NSK Dental, Japan).
Following the placement of viral mixtures into the quartz capillary pipette, the pipette was slowly (0.2 mm/s) maneuvered into the desired location within the brain tissue. Using bregma as a reference point, the coordinates of anterio-posterior (AP): 3.3mm, dorso-ventral (DV): 2.1mm, and medio-lateral (ML): 0.7mm and 1.2mm for targeting medial and lateral regions of the superior colliculus. For the medial inferior olive: AP: 6.1mm, ML: 0.4mm, DV: 6.72mm and for the principal olive: AP: 6.4mm, ML: 0.2mm, DV: 6.72mm. At these locations, 20–40 nl of viral mixtures were injected with a speed of 20 nl/min. Approximately 5–10 min after the completion of the viral delivery, the pipette was retrieved from the tissue with slow speed. The skin was cleaned and the incision was sutured, followed by subcutaneous administration of 5 mg/kg of Rymadil (Zoetis, NJ, USA) to minimize the post-operative discomfort. Following stereotaxic viral injections, mice were placed on a heating pad and monitored continuously until they regained full mobility. Mice were returned to their cage with a familiar cage mate to reduce stress. Health status, grooming, and overall activity were monitored daily for at least 3 days post-surgery. Animals showing signs of distress, infection, or abnormal behavior were promptly evaluated and treated in accordance with institutional animal care protocols.
Viral vectors
For anterograde labeling of superior colliculus neurons (SC), we use AAV9.hSyn.eGFP (final titer: \(7 \times 10^\) vg/ml, 50465, Addgene) and a mixture of AAV9.hSyn.Cre (105553, Addgene) or AAV9.CamKII.Cre (105558, Addgene) with AAV9.CAG.Flex.tdTomato (28306, Addgene). These viral vectors were carefully mixed and subsequently diluted in phosphate buffered saline (PBS) to achieve a final titer of \(1.7 \times 10^\) vg/ml.
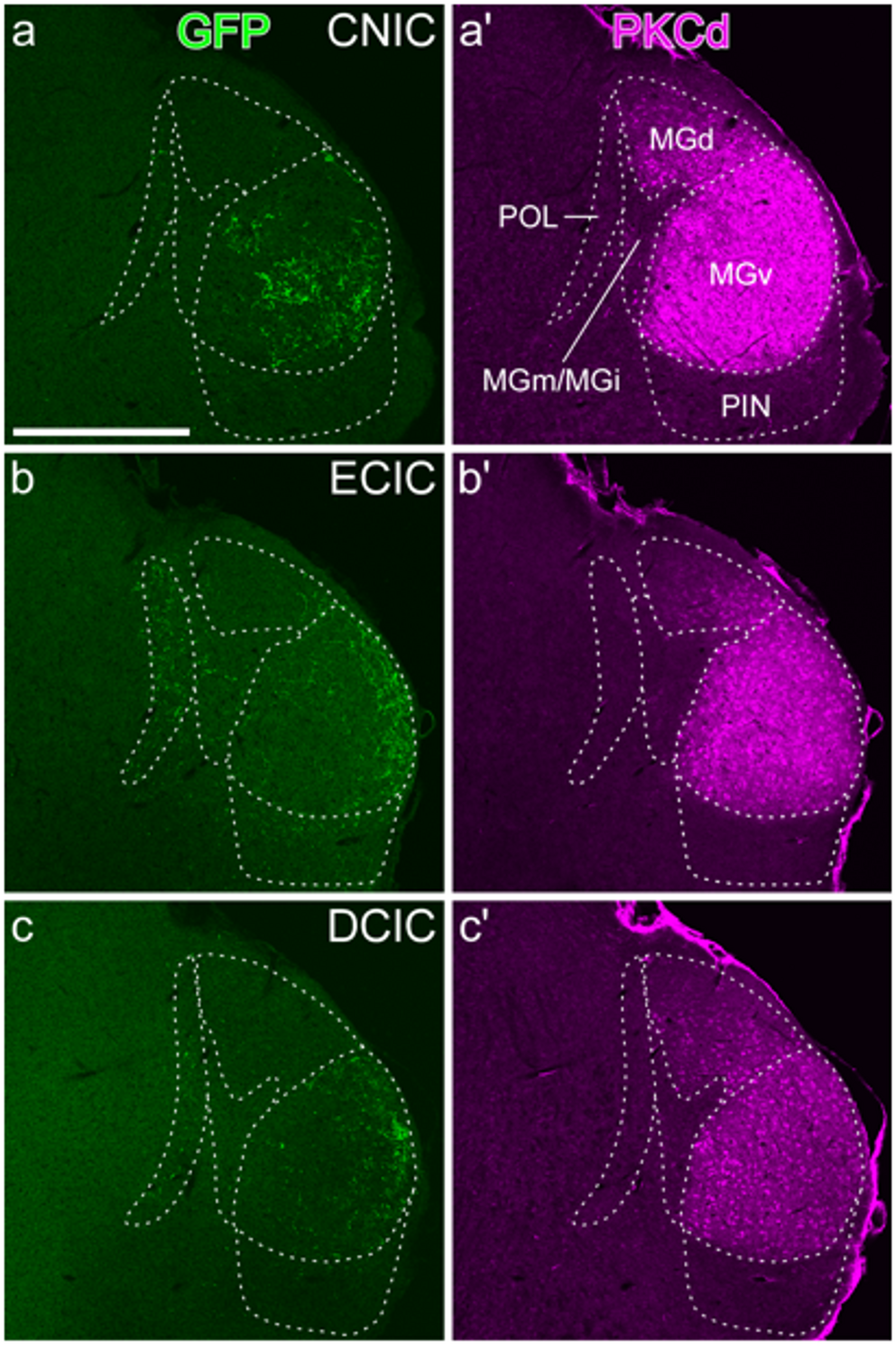
To retrogradely label the afferents in different inferior olive subnuclei (IO), we used AAVrg.CAG.tdTomato (final titer: \(3.5 \times 10^\) vg/mL, 59462, Addgene) and AAVrg.CAG.eGFP (final titer: \(3.5 \times 10^\) vg/mL, 37825, Addgene) in the medial accessory olive (MAO) and principal olive (PO).
The AAV9.hSyn.Cre and AAV9.CAG.Flex.tdTomato mixture, along with AAV.PHP.S.Htr5b.eGFP (final titer: \(5 \times 10^\) vg/mL, OIST), was used to label SC axons and sparsely label IO neurons to identify the location of SC synapses along the IO. For in vivo calcium imaging of IO neurons, we used a mixture of IO-specific viruses: AAV.PHP.eB.TRE.GCaMP6s (final titer: \(2 \times 10^\) vg/mL, OIST) and AAV.PHP.eB.Htr5b.tTA (final titer: \(2 \times 10^\) vg/mL, OIST Dorgans et al. (2022)). Furthermore, AAV9.Syn.ChrimsonR.tdTomato was used for optogenetic activation of SC axons.
Tissue processing and immunohistochemistry
Two to three weeks after injection of viral tracers into the superior colliculus (SC), the animals were transcardically perfused with PBS followed by 4% PFA (w/v, in PBS). The brains were extracted and postfixed in 4% PFA solution for 2–3 h at room temperature, after which they were stored in PBS at 4 \(^\) C until sectioning. The brains were sectioned using a vibratome (Model 5100MZ-plus, Campden Instruments, Loughborough, UK) equipped with ceramic blades (Model 7550–1-C, 38 x 7 x 0.5 mm, Campden Instruments, Loughborough, UK). The slices were mounted with Vectashield (Vector laboratories, CA, USA) or Prolong Glass (Invitrogen, MA, USA) and #1.5 coverslip glass (Harvard Apparatus, MA). For immunohistochemistry, the brains were sectioned either with with the vibratome or with a cryostat (CM1950, Leica, IL, USA) equipped with blades (Disposable Blades DB80LX, Leica, IL, USA). The brains that were sectioned with the cryostat were cryoprotected by submerging them in a 10% sucrose solution (w/v, in PBS) for several hours at room temperature, followed by being stored overnight in a 30% sucrose solution (w/v, in PBS) before sectioning. To verify injection sites in the SC, the rostral parts of the brains were sectioned coronally at 50 \(\mu\)m or 100 \(\mu\)m. These sections were washed 2x in PBS for 10 min at room temperature, followed by 3 X 5 min washes in 0.1M phosphate buffer (PB) at room temperature, after which they were mounted on an objective glass with Prolong glass mounting medium (Invitrogen, MA, USA) and #1.5 coverslip glass (Harvard Apparatus, MA, USA).
The brainstems were sectioned separately at 50 \(\mu\)m in coronal orientation, also using the vibratome with ceramic blades or cryostat. For immunostaining, every second brainstem section was stained freely-floating using anti-VGluT2 (Polyclonal, Thermo Fisher, MA, USA) and anti-VGAT (Polyclonal, Alomone labs, Jerusalem, Israel). The sections were first washed 4 times in PBS at room temperature for at least 10 min. Subsequently, they were blocked and permeabilized in PBS containing 10% Normal Goat Serum (NGS, Abcam, Cambridge, UK) and 0.5% Triton X (Thermo Fisher, MA, USA) for 2 h on a shaker (100 rpm), followed by primary antibody incubation. For this step, the slices were incubated for 48 to 72 h at 4\(^\)C in a solution of PBS with 2% NGS and 0.1% Triton X and anti-VGluT2 (1:500) and anti-VGAT (1:2000) primary antibodies. The sections were subsequently washed 5 times for 10 min in PBS at room temperature, before secondary antibody incubation for 2 h at room temperature, in a PBS solution with 2% NGS, 0. 1% Triton X and secondary antibodies. The secondary antibodies used were goat-anti rabbit-A647 (Invitrogen, MA, USA) and goat-anti guinea pig-A555 (Invitrogen, MA, USA), both at a concentration of 1:2000. Finally, sections were washed 2 times for 10 min in PBS, followed by 3 washes in 0.1M phosphate buffer (PB), before being mounted using the same procedure as viral-labeled sections.
Confocal image acquisition and image processing
Z-stack images of sections labeled using viral and immunohistochemical methods were acquired using a Zeiss LSM 880 confocal system (Carl Zeiss Microscopy, Jena, Germany). For overview scanning of the entire cortex and brainstem, a 10x objective (Plan-Apochromat 10x, NA = 0.5; Carl Zeiss Microscopy, Jena, Germany) with a 6 \(\mu\)m z-step was used. To scan the inferior olive (IO), a 20xobjective (Plan-Apochromat 20x, NA = 0.8; Carl Zeiss Microscopy, Jena, Germany) with a 3–6 \(\mu\)m z-step was used. For the superior colliculus (SC), a 10x objective (Plan-Apochromat 10x, NA = 0.5; Carl Zeiss Microscopy, Jena, Germany) with a 6 \(\mu\)m z-step was used. Higher-magnification images were acquired using a 40x oil-immersion objective (Plan-Apochromat 40x Oil DIC M27, NA = 1.4; Zeiss Immersol oil; Carl Zeiss Microscopy, Jena, Germany) with 0.07\(-\)0.1 \(\mu\)m z-steps, adjusted to match the x and y resolution. All images were acquired at 1024x1024 pixel resolution.
For multichannel imaging, the following excitation/emission settings were used: 488 nm/490–535 nm for eGFP, 561 nm/470–655 nm for tdTomato, and 633 nm/638–758 nm for the infrared channel. Acquisition parameters were optimized for each experiment but kept constant across samples within the same experimental group. After acquisition, z-stacks were processed using a 3D Gaussian filter with \(\sigma\) = 1 pixel in x, y, and z dimensions, and converted into 2D images using a standard deviation projection along the z-axis.
Percentage of SC-IO axons labeling across IO subnuclei
After acquiring images from the IO, we classified them into three rostro-caudal regions: caudal IO, middle IO, and rostral IO. The subnuclei within each IO region were manually delineated using Fiji (Image J, DC, USA). We classified the subnucleus a, b and c of the inferior olive (IOA, IOB, and IOC) as ventral MAO and classified IOVL (inferior olive, ventrolateral protrusion), IOBe (inferior olive, beta subnucleus), IOK (inferior olive, cap of Kooy), and IODM (inferior olive, dorsomedial) as dorsal MAO.
For each rostro-caudal IO region, we quantified the intensity of labeled SC axons in different IO subnuclei using slices from five animals. Intensity values were normalized to the maximum intensity value for each animal (see Supplementary Table 1) to generate the values presented in Fig. 1H1. Normalized values below 0.2 were considered indicative of passing axons or background noise from image acquisition.
Soma areas and position of the SC-IO neurons
To locate the labeled SC-IO neurons along the rostro-caudal axis, the midline position of the acquired confocal images of the midbrain was manually aligned and the coordinates of each slice were matched to the bregma position using the Mouse Brain Atlas (Paxinos and Franklin 2001). The centers of mass of the soma boundaries were used as x-y positions, and their distance to the midline and SC surface was calculated. To further map the labeled neurons along the mediolateral axis (M-L), the locations of the soma were transformed into ’angular space’, following the conventions described in Benavidez et al. (2021).
Neuronal tracing and identification of the SC-IO putative synaptic contact
To identify the location of synapses, processed confocal images of IO neurons and SC axons were imported into Vaa3D (Peng et al. 2014), an open-source platform for high-resolution 3D visualization and neuron tracing. IO neurons and SC axons were manually traced in three dimensions, and points of close apposition were identified as putative synapses. The number of synapses was quantified and exported to Excel and each contact was manually classified as either spine- or shaft-contact. The thickness of IO dendrites at the sites of putative SC-MAO and SC-PO synapses was manually measured.
Next, to quantify the ratio of VGluT2+ to VGAT+ boutons in SC fibers, we identified several 100 x 100 \(\mu\)m regions of interest within the caudal medial accessory olive (cdMAO) and the principal olive (PO) eGFP+ SC axons in immunolabeled slices. We selected ROIs per animal (two ROIs within the MAO and one ROI in the ventral PO, on both sides).
Z-stacks of these ROIs were acquired using a confocal microscope with a 40x objective. The images are further divided into four subregions to facilitate analysis. In each of the subregions, we applied noise reduction using the 3D Gaussian filter (\(\sigma\) x,y,z =1 pixel) and manually quantified the percentage of GFP+ boutons labeled with VGluT2, VGAT, both and none, using the following procedures in the FIJI software. Afterwards, the analysis was performed in two steps using custom FIJI macros.
The first step aimed to identify eGFP+ boutons (indicating that the origin of the axon was in the SC). For the PO, the number of boutons was rather low, allowing us to identify and label all boutons in the image. This was done by manually drawing ROIs in FIJI using the free-hand or polygon selection tool. For densely labeled boutons in MAO it was not feasible to label all boutons individually. Instead, a random subset of boutons was selected. This selection process was performed manually with the assistance of a TANGO macro (Ollion et al. 2013). The macro identified local maxima that exceeded an intensity threshold of 2500 using a local maximum filter from the ImageJ 3DSuite package. The macro then randomly presented to the person analyzing (observer) with one of the identified local maxima, allowing them to draw an ROI if the local maximum was deemed to correspond to a bouton. This process continued until all local maxima in the image were evaluated or a predetermined threshold (set at 50 boutons) was reached for the subregion.
The second step aimed to determine whether the boutons identified in step one were labeled with VGluT2 or VGAT. A macro presented each ROI to the user for manual categorization as labeled, unlabeled, or doubtful for both the VGluT2 and VGAT channels. For analysis purposes, boutons marked as "doubtful" were treated as unlabeled. Furthermore, users could exclude any bouton from the analysis if they were not completely confident that ROI represented a synaptic bouton. To avoid subjective bias, each step was analyzed by different observers. For example, if step one was conducted by observer A, step two was performed by observer B.
In-vivo calcium imaging of IO neuronal population and pptogenetic activation of SC axons
For calcium imaging experiments, we used a ventral access surgery. The procedure is conducted under deep anesthesia (isofluorane 1.5–2%) and the animal is not allowed to regain consciousness after the recording is completed.
Two to three weeks after injecting AAV9.Syn.ChrimsonR.tdTomato into the superior colliculus (SC) and AAV.PHPeB.TRE.GCaMP6s along with AAV.PHPeB.Htr5b.tTA into the inferior olive (IO), mice were anesthetized with 5% isoflurane for ventral surgery to allow placement of the GRIN lens (Guo et al. 2021). Prior to surgery, the thigh and throat were shaved, with thigh sensors used to monitor oxygen and breathing rate, and a rectal probe used to monitor body temperature.
An incision was made on the throat after the application of lidocaine. The salivary glands were displaced and the sternothyroid muscles overlying the trachea were removed for tracheotomy. The thyroid gland was removed and a thread was secured around the tracheal ring to stabilize it before connecting it to a pre-made intubation tube, ensuring the maintenance of the anesthesia. The esophagus, larynx, and surrounding tissues were removed, exposing the occipital bones, ventral arch, and anterior tubercle of the atlas. The ventral arches of the atlas and anterior tubercle were removed using a rongeur, and the surrounding occipital bone was cut to enlarge the field of view. The cartilage overlying the dura was removed to expose the ventral IO to have a clear view during calcium recording.
A GRIN lens (9 mm length, 1 mm diameter) connected to an Inscopix mini-microscope was placed on the ventral IO surface. For the GCaMP6s recording, blue LED light (455 ± 8 nm) was used, while red light (620 ± 30 nm) was used for optogenetic stimulation. The 40 Hz recordings lasted for 120 s each. For optogenetic stimulation, a 100-ms pulse train (20 Hz, 5 ms pulses) was delivered 30 s after the recording began, with nine repetitions with a 10-second interval between the stimulations. The recordings were spatially filtered and motion corrected using Inscopix Data Processing (IDP) software. The regions of interest (ROIs) were manually traced, and the fluorescence traces were analyzed and visualized in MATLAB.
Data analysis and statistics
For the analysis of SC-IO neuron coordinates and soma size, all detectable SC-PO and SC-MAO neurons were manually delineated in Fiji following image processing, and saved as Regions of Interest (ROIs). Neurons with poorly defined soma boundaries or very faint fluorescence were excluded. Each ROI was classified as SC-MAO, SC-PO, or co-labeled (both), and soma area was subsequently measured. Measurements were imported into GraphPad Prism for data visualization and statistical analysis. Values are reported as mean ± SEM.
To compare cumulative distributions of SC-IO, non–SC-IO, SC-MAO, and SC-PO neurons, plots were generated using custom MATLAB code, and statistical comparisons were performed using the Kolmogorov–Smirnov test. The spatial distributions of SC-MAO and SC-PO neurons along the rostro-caudal and medio-lateral axes were also visualized using custom-written MATLAB scripts.
IHC analysis was assisted by a custom FIJI macro adapted from a previously published tool (Ollion et al. 2013). Results were recorded and summarized in a Microsoft Excel file, and data visualization was performed in GraphPad Prism. For synapse localization analysis, GraphPad Prism was used to visualize the distribution of synapse positions and comparison of the spine vs shaft contact.
Following spatial filtering and motion correction, all subsequent analysis and visualization of the calcium imaging data were performed using custom-written MATLAB code. Fluorescence traces were low-pass filtered and aligned to stimulus onset for visualization. Cross-correlation matrices were generated in MATLAB, along with the mean cross-correlation value between trials.



Comments (0)