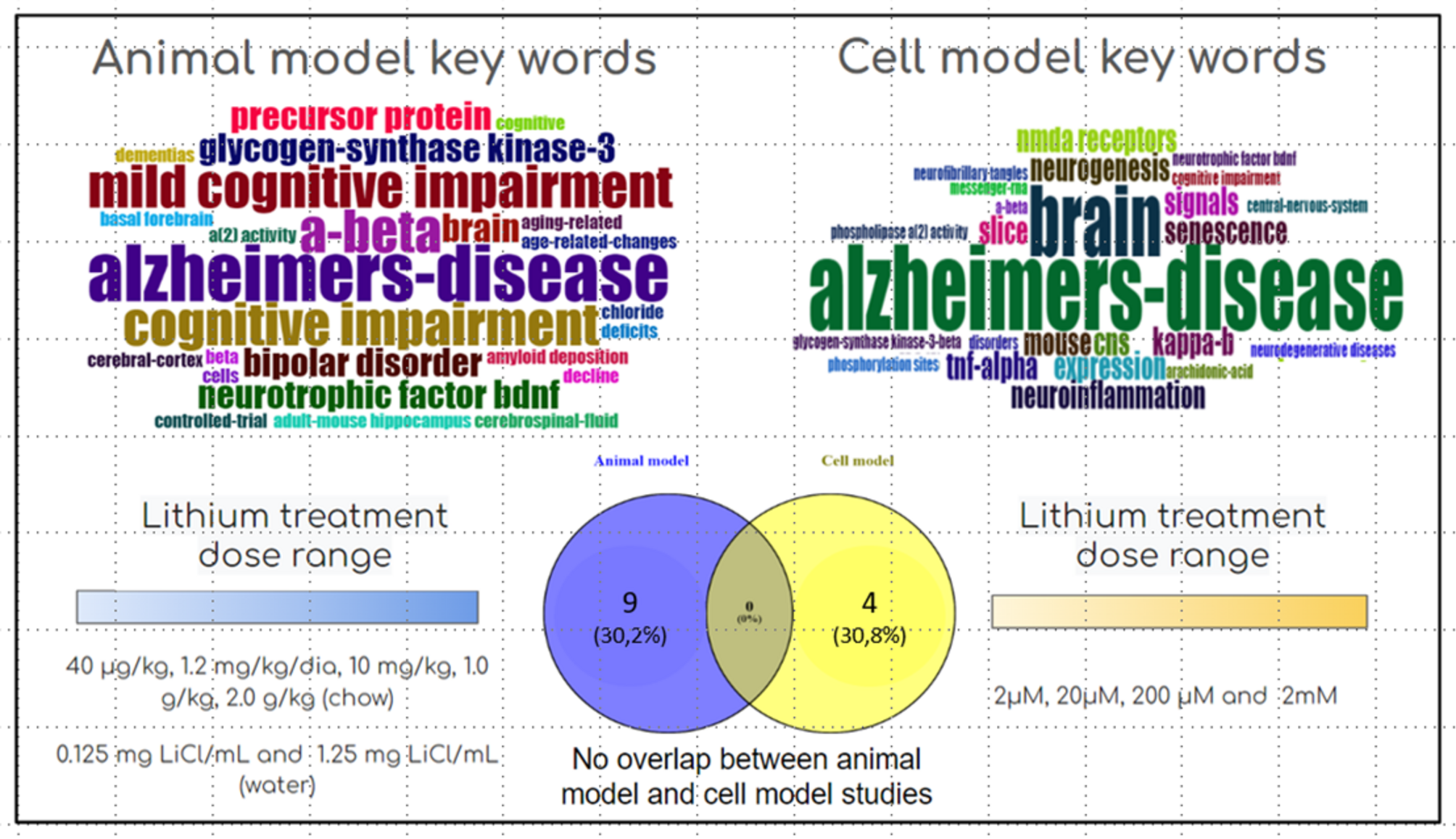
Animal models
Animal models are widely used in investigating AD, yielding critical insights into the pathology, pharmacology, and behavioral changes associated with the condition (Quiroz et al. 2010; Methaneethorn 2018). Effectively translating findings from these models into clinical applications for humans requires a meticulous approach that considers the specific characteristics of each model. In rodents, lithium is rapidly absorbed from the gastrointestinal tract, distributed throughout total body water without binding to plasma proteins, and is almost exclusively eliminated by the kidneys (Wood et al. 1986; Wraae 1978). In rats, lithium serum concentrations peak approximately one hour after the administration of a 2 mmol/kg dose (Ebadi et al. 1974). In the brain, peak concentrations occur within 4 to 6 h (Wraae 1978), corresponding to approximately 40% of peak serum levels (Mukherjee et al. 1976). The half-life of lithium in serum ranges from 8 to 10 h, while in the brain, it extends to 24 to 36 h (Ebadi et al. 1974; Martinsson et al. 2013; Mukherjee et al. 1976). These pharmacokinetic differences highlight the importance of dose adjustments and considering species-specific characteristics to accurately extrapolate preclinical findings to clinical practice (Wraae 1978).
Mury et al. (2016) demonstrated that chronic administration of lithium at a dose of 2 mmol/kg increased phospholipase A2 (PLA2) activity in the brain, particularly in the cortex and hippocampus. The activation of brain PLA2, an enzyme critical for lipid signaling and membrane remodeling, constitutes a key mechanism through which lithium enhances memory performance in rodent models. This finding suggests that subtherapeutic doses of lithium promote the production of bioactive lipids that directly facilitate synaptic plasticity and cognitive function.
Glycogen synthase kinase-3 beta (GSK3β), a central enzyme involved in tau phosphorylation and amyloid precursor protein (APP) processing, is another pivotal target modulated by lithium. Subtherapeutic doses of lithium (0.2 mmol/kg/day) decrease GSK3β activity in key brain regions, including the hippocampus and prefrontal cortex (Fenech et al. 2023). This reduction in GSK3β activity directly correlates with diminished tau hyperphosphorylation and plays a crucial role in attenuating amyloidogenic APP processing—both processes being critical hallmarks of AD pathology. Furthermore, long-term administration of lithium (2 mmol/kg/day for 4 months) has been shown to preserve the CA1 pyramidal cell layer in 3xTg-AD mice by decreasing neuronal loss (Schaeffer et al. 2017). The neuroprotective pathway is similarly effective in maintaining the integrity of the CA1 pyramidal cell layer, thereby reducing cell loss. Structural preservation of the hippocampus, a region essential for memory and learning, has also been documented with subtherapeutic lithium treatment. In an obesity model, lithium supplementation has been observed to decrease GSK3 activity and enhance insulin sensitivity, indicating potential broader metabolic implications (Fenech et al. 2023).
Lithium administration has been shown to reduce markers associated with oxidative stress, thereby contributing to its neuroprotective effects. Additionally, varying doses of lithium treatment have decreased oxidative stress markers, further supporting these neuroprotective outcomes (Schaeffer et al. 2017). Lithium increases the levels of calcium/calmodulin-dependent protein kinase II (CaMKII) and BDNF, both of which are critical for memory formation, neuronal survival, and synaptic resilience. Beyond its biological benefits, the impact of lithium on behavioral and cognitive outcomes is substantial. Notably, Nunes et al. (2015) found that long-term administration of subtherapeutic doses of lithium (0.25 mmol/kg/day over 8 weeks) resulted in the reduction of neurofibrillary tangles and amyloid plaques in a transgenic mouse model of AD.
In the context of amyloid pathology, Wilson et al. (2017) demonstrated that the administration of a subtherapeutic dose of lithium (NP03) at 0.2 mmol/kg/day for 8 weeks in McGill-R-Thy1-APP transgenic rats resulted in a significant reduction in the activity of the APP-cleaving enzyme 1 (BACE1). This decrease in BACE1 activity correlated with lower levels of early-stage amyloid pathology and improved memory function in transgenic mice. This suggests that lithium effectively targets upstream mechanisms involved in AD progression. Furthermore, the therapeutic benefits of lithium extend beyond amyloid plaques and tau-related pathology. Subsequently, Wilson et al. (2020) investigated the effects of similar doses of NP03 in the same mouse model during a 12-week trial that covered the transition phase from Aβ plaque-free to plaque-formation, reporting a reduced loss of cholinergic boutons in the hippocampus, lower levels of soluble and insoluble cortical Aβ42, decreased hippocampal Aβ plaque deposition, and diminished evidence of neuroinflammation and cellular oxidative stress, all coupled with improvements in memory function.
Pereira et al. (2024) conducted a study using SAMP-8 mice, a model representative of accelerated aging, in which a similar subtherapeutic dosage of 0.2 mmol/kg/day was administered over 8 weeks. The findings revealed that lithium treatment reduced neuroinflammatory markers, such as interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α), while simultaneously elevating levels of synaptic proteins, including synaptophysin, which indicates enhanced synaptic functionality. These results underscore lithium’s potential to ameliorate neuroinflammation and promote synaptic health.
Lithium also enhances and regulates neuronal-glial interactions, which are essential for sustaining neuronal viability and synaptic health. Rocha et al. (2020) found that subtherapeutic lithium improves astrocytic and microglial functions, as well as neuronal communication. Furthermore, the effect of subtherapeutic lithium on senescence and cellular aging has been demonstrated by its capacity to preserve telomere length. Cardillo et al. (2018) reported that chronic lithium treatment (2 mmol/kg/day for 6 months) resulted in increased telomere length in the parietal cortex and hippocampus of 3xTg-AD mice, which was associated with reduced markers of cellular senescence and improved genomic stability. These studies underscore lithium’s significant role as a neuroprotective agent capable of modulating critical molecular pathways, maintaining brain structure, and enhancing cognitive function. Its influence on enzymes such as GSK3β and BACE1, along with its potential to augment neurotrophic signaling and mitigate neuroinflammation, highlights its therapeutic promise in AD. Nevertheless, further research is essential to refine dosing strategies, evaluate long-term safety, and effectively translate these findings into clinical applications.
Cellular models
Cellular models are crucial instruments in studying the CNS. They afford comprehensive insights into the intercellular communication and functionality of neuronal cells. These models are especially significant in investigating neuroprotective mechanisms and neurodegenerative pathways, enhancing our understanding of the underlying biological processes.
In organotypic cultures of the hippocampus derived from SAMP-8 mice, very low doses of lithium demonstrated a crucial role in neuroprotection, with reduced neuronal loss and neuroinflammation in older individuals. These low doses also promoted a significant reduction in the CA2 area of the hippocampus and reduced the activation of NFkB and the density of inflammatory cytokines (Toricelli et al. 2021). In primary cultures of cortical and hippocampal neurons, prolonged exposure to lithium (0.02 mM, 0.2 mM, and 2 mM over 7 days) resulted in increased metabolism of membrane phospholipids, with activation of total phospholipase A2, c- and iPLA2 (De-Paula et al., 2015). Co-cultures of cortical and hippocampal neurons with glial cells showed that chronic treatment with lithium at subtherapeutic concentrations modified the secretion of pro- and anti-inflammatory interleukins (De-Paula et al. 2016a, b). Chronic treatment with lithium at subtherapeutic doses also increased cell viability in primary cultures of cortical and hippocampal neurons. It mitigated the shortening of neuronal telomeres despite the presence of induced amyloid toxicity (Themoteo et al. 2022). Finally, De-Paula and Forlenza (2020) also investigated the effects of chronic treatment with lithium at different concentrations (including subtherapeutic doses) in primary cultures of cortical and hippocampal neurons, showing that lithium modulates the expression of several tau kinases, such as GSK3β, Akt, PKA, and CaMKII, with distinct responses between cortical and hippocampal neurons. Hippocampal neurons showed greater sensitivity to lithium at lower doses, while significant changes in cortical neurons occurred only at therapeutic doses, highlighting the regional specificity of lithium’s effects. These findings underscore that lithium, even at low concentrations, can confer substantial benefits regarding neuroprotection and the prevention of neurodegeneration. That emphasizes the necessity for further investigations to elucidate its therapeutic potential and long-term safety.






Comments (0)