Anxiety, when experienced in response to threat, serves as an evolutionarily adaptive mechanism that promotes vigilance and facilitates appropriate and adaptive behavioral responses. However, in some individuals, anxiety can become maladaptive, marked by heightened physiological and emotional reactivity that interferes with daily functioning [1]. Understanding the processes that underlie this transition from adaptive to dysfunctional anxiety is crucial for characterizing the dimensionality of its experience and developing effective interventions. Although some mechanisms are already known, current pharmacological treatments often lack specificity and show high variability in effectiveness [2]. As Ren et al. emphasize [3], despite the availability of a range of treatments, nearly 40% of individuals with anxiety disorders fail to respond to one or more medications. This underscores the need for deeper insights into the different underlying processes and neurobiological mechanisms of anxiety, beyond what is captured by diagnostic categories.
One of the central components of the stress response that has been widely studied in this context is the hypothalamic-pituitary-adrenal (HPA) axis. This neuroendocrine system regulates the secretion of cortisol, a glucocorticoid hormone essential for maintaining physiological homeostasis during environmental challenges [4, 5]. In response to acute stress, the HPA axis (along with the sympathetic nervous system) mobilizes bodily and cognitive resources, increasing heart rate, blood pressure, and cortisol levels to support adaptive action [6]. However, under conditions of chronic or intense stress, the HPA axis can become dysregulated, disrupting negative feedback mechanisms and leading to sustained cortisol secretion [7]. This dysregulation has cascading effects on multiple systems, including metabolism, immune function, mood, and cognition [8]. Thus, while cortisol is vital for short-term adaptation, persistent elevations have been linked to maladaptive outcomes and increased risk for psychiatric conditions such as anxiety and mood disorders [5, 9]. Chronically elevated cortisol disrupts activity in areas rich in glucocorticoid receptors, particularly the prefrontal cortex (PFC), which governs executive function and top-down control of emotion [10]. Concurrently, elevated cortisol increases activation in limbic and subcortical regions, such as the amygdala, contributing to emotional hyperreactivity [11,12,13]. These patterns highlight the role of the HPA axis in modulating prefrontal-limbic circuitry essential for emotional regulation [6].
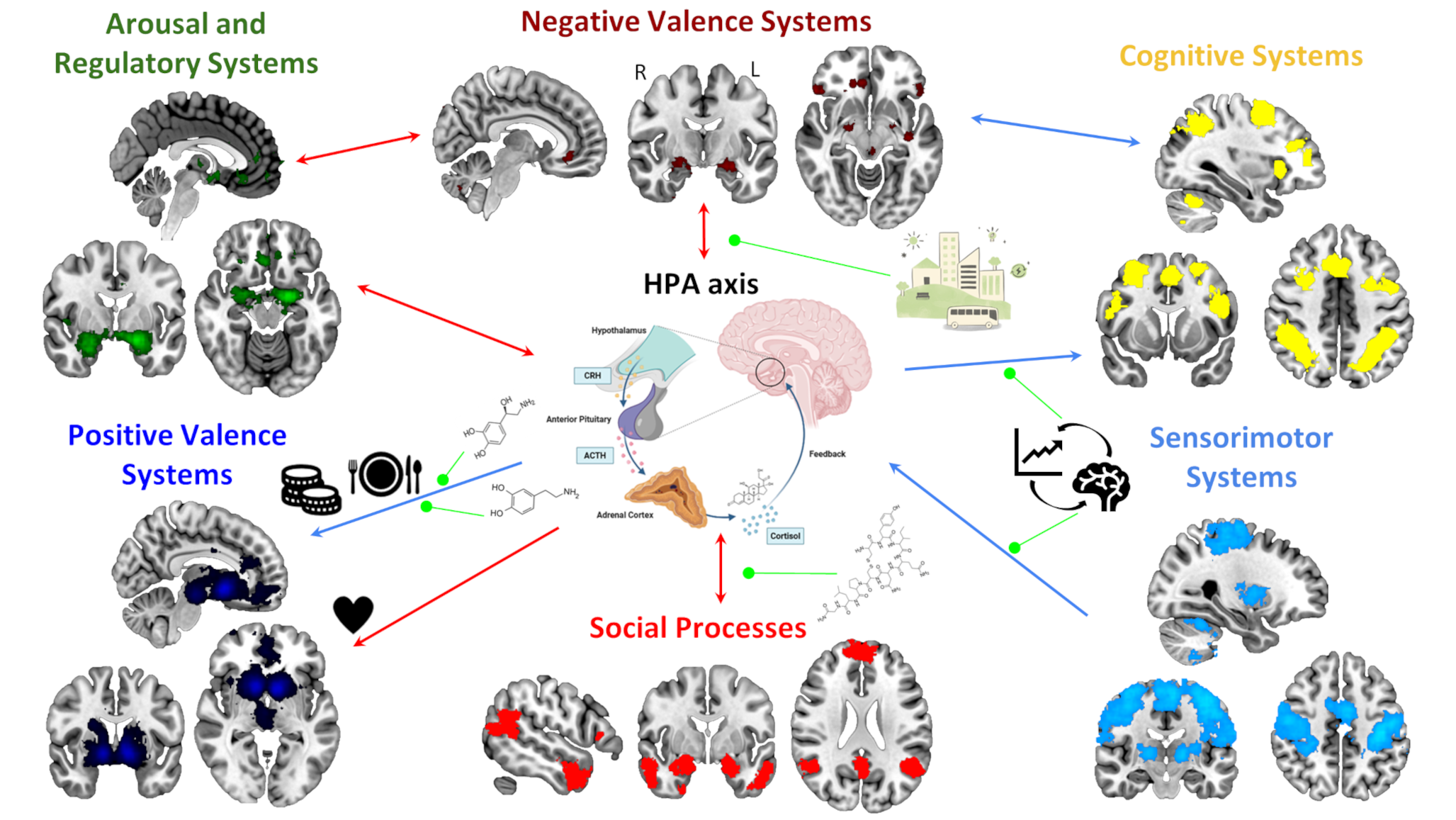
Despite considerable progress in understanding these mechanisms, most research to date has focused on a categorical diagnostic framework. This limits our understanding of how stress-related physiological dysregulation, such as altered cortisol patterns, manifests in subclinical, at-risk, or comorbid populations. As Bate et al. note [14], a dimensional approach may be better suited to capture the non-linear and heterogeneous associations between cortisol levels and symptom severity across a broader spectrum of functioning. The Research Domain Criteria (RDoC) framework, developed by the National Institute of Mental Health, offers a transdiagnostic, multi-level model for investigating mental health beyond traditional diagnostic boundaries. RDoC conceptualizes mental functioning across six major domains: Negative Valence Systems, Positive Valence Systems, Cognitive Systems, Social Processes, Arousal and Regulatory Systems, and Sensorimotor Systems. These are evaluated across multiple units of analysis (e.g., genes, circuits, behavior) and can vary in severity from normative to pathological [15, 16].
As Rief et al. argue [17], mental health is shaped by a dynamic interaction of biological, psychological, social, and environmental factors. Individuals with the same diagnosis may differ in the underlying causes and symptoms they present. Moreover, symptoms themselves are often causally interconnected and fluctuate over time. For example, individuals with high levels of anxiety may experience overlapping emotional, cognitive, and physiological disturbances that reinforce one another. These patterns challenge simplistic cause-effect models and underscore the need for more flexible, dimensional frameworks that capture mental health at varying levels of analysis and degrees of impairment.
HPA axis dysregulation and cortisol dynamics align closely with RDoC constructs, particularly within the Negative Valence and Arousal and Regulatory Systems domains, making them promising candidates for exploring stress-related vulnerability across diagnostic boundaries. Furthermore, the RDoC framework supports an integrated approach to identifying cross-cutting symptom profiles and mapping them onto underlying biological and behavioral processes. By integrating molecular, neural, and behavioral data, this framework facilitates investigation of how stress reactivity contributes not only to clinical anxiety but also to broader patterns of emotional and cognitive dysregulation. This is, specific processes that, if dysregulated, can be related to high levels of anxiety, its onset and maintenance, and even worsening.
Although research using the RDoC approach has grown in recent years, comprehensive reviews linking stress biomarkers (such as cortisol) to anxiety-related processes remain limited. Addressing this gap could clarify the role of HPA axis dysfunction in stress and affective disruptions and inform the future development of the RDoC framework itself, which is designed to evolve as scientific evidence accumulates [15, 17]. Thus, in the coming sections we will provide an overview of studies exploring the association between the HPA axis and anxiety-related processes (accompanied by their neural correlates, when available), for each of the six major RDoC domains. At times, we will discuss processes that, while not explicitly labeled as anxiety-related in the original studies, are closely linked to anxiety. In other instances, we will focus on studies that directly investigate anxiety-related phenomena. This approach underscores the importance of studying these processes dimensionally, while also offering attention to how shared mechanisms may underlie diverse mental health conditions.
Negative Valence Systems
Negative Valence Systems have been widely investigated to understand mental health conditions such as anxiety disorders, depression and post-traumatic stress disorder (PTSD). Aversive stimuli lead to neural responses in limbic and limbic-association regions accompanied by physiological changes such as increased cortisol levels and heart rate. We aim to describe these processes, along with the atypical patterns observed in mental health disorders, focusing on three key domains: threat (acute and sustained), grief, and frustrative non-reward.
Threat can lead to fear and anxiety, where fear is defined as a phasic response to certain-and-imminent danger, whereas “anxiety” is a sustained response to uncertain-or-distal harm [18, 19]. The most common response to imminent danger is the freeze response. In contrast to the classical view, Roelofs and Dayan propose that freezing does not reflect a passive fear state [19]. Rather, it involves the active coordination of cognitive and somatic responses following a threat encounter, allowing the body and mind to prepare for action. These processes are mediated by the neurotransmitters acetylcholine (ACh) and noradrenaline (NA). NA is produced by the locus coeruleus and influences cognitive function such as arousal, attention and perception. In contrast, ACh is responsible for parasympathetic nervous system activity such as reduced motion, heart rate and breathing. As such, freezing is associated with concurrent sympathetic and parasympathetic upregulation, with the latter being dominant. This reflects a state of poised readiness rather than passive withdrawal.
Sustained threat also triggers hypervigilance and hyperarousal, mediated by the bed nucleus of the stria terminalis (BNST). Atypical BNST activity and functional connectivity (FC) to the amygdala have been linked to neurodevelopmental traits in social anxiety disorder (SAD) [20], youth exposure to interpersonal violence [21] and PTSD in veterans [22]. Furthermore, social threat, induced by performing a challenging task under observation, activates the amygdala, anterior cingulate cortex (ACC), insula and thalamus, involved in the detection of salient stimuli and threats [23, 24]. Moreover, aberrant FC within the salience and limbic networks is associated with anxiety [25] and PTSD [26, 27], highlighting their role in mental health. Environmental context also matters: well-connected urban designs are linked to reduced activity in these stress-responsive brain regions [23], highlighting the potential of urban planning and social infrastructure to buffer against psychological distress. Social evaluative stress also elevates HPA activity, especially in anxiety disorders. However, Labuschagne et al. emphasize the need for standardised cortisol measures, allowing for comparisons across studies [28]. This is important as this research forms the basis for medical clinical trials on the use of cortisol-based medicine for anxiety disorders and PTSD [29].
Grief following loss also elicits negative emotional responses, reflected by neural and physiological changes. Early after suicide loss, individuals show attentional bias toward deceased-related cues, linked to increased activity in the dorsolateral (dlPFC) and ventrolateral PFC (vlPFC), the orbitofrontal cortex (OFC), and the insula. Reduced engagement of these areas is associated with greater avoidance of reminders, suggesting that weaker cognitive control may lead to maladaptive coping [30]. Furthermore, striatal connectivity with emotional and cognitive control regions is influenced by grief intensity and yearning in the elderly. Blair et al. suggest that dysfunction of the striatal reward system may contribute to prolonged grief disorder [31], though more research is needed. Grief-like responses also arise during parent-child separation. Separation anxiety influences the HPA axis, which in turn influences the autonomic nervous system and the immune-inflammatory system. The latter is associated with depressive processes and this could explain learned helplessness in adolescents with separation anxiety [32]. Notably, age of separation is a key moderating factor [33] (see Table 1).
Table 1 Overview of studies looking at the influence of the HPA axis on each of the six major RDoC domainsFrustrative nonreward is defined as the psychological state induced when an expected reward is withheld. Irritability shares a negative valence with anxiety and is a significant predictor of its development [34]. Children with high levels of irritability have greater frontal-striatal activation with induced frustration [35]. Furthermore, FC during frustration predicts child-reported irritability as shown by connectome-based predictive modelling [36]. In addition, previous studies suggest that variations in HPA axis functioning can influence the developmental trajectory of irritability and internalizing symptoms [37, 38] (Table 1).
Positive Valence Systems
Positive Valence Systems are responsible for responses to positive motivational situations or contexts, such as reward seeking, consummatory behavior, and reward/habit learning. At the brain level, reward processing networks are composed of the striatum, the amygdala, the OFC, and the ventromedial PFC (vmPFC)/ACC [39]. Moreover, these systems are thought to be influenced by cortisol reactivity. Stress-induced changes in HPA axis functioning have been shown to be related to reductions in the neural correlates of reward processing during reward tasks [40], an effect that can be reversed by methylphenidate-induced increases in dopamine and NA [41]. A review by Harrewijn et al. had similar findings, with reduced activity in the amygdala during reward tasks after exogenous cortisol administration [42]. This goes in line with studies on emotional eaters [43], and exploring the effect of hydrocortisone on hunger [44], while the opposite effect has been found for sex stimuli – that is, higher cortisol associated with an increase in approach behavior and activation in reward processing areas [45, 46] (Table 1).
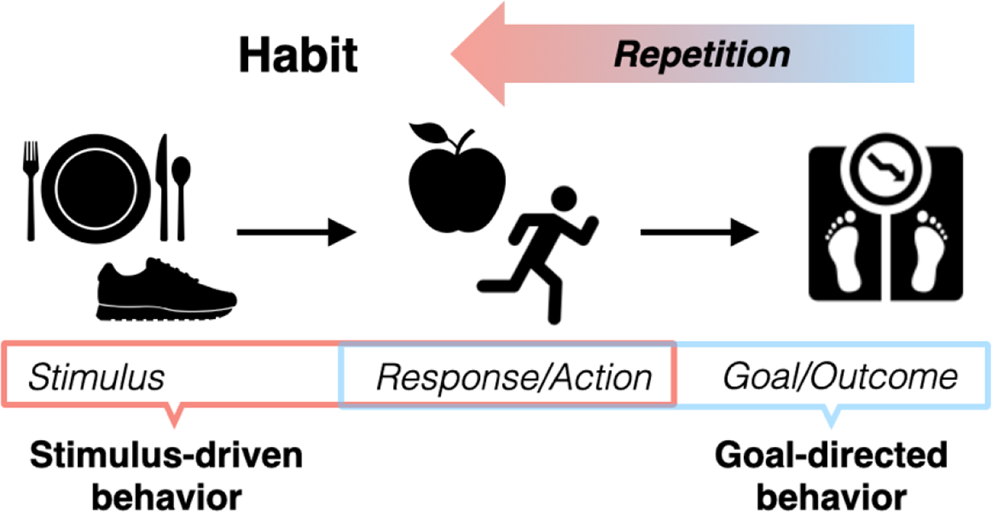
Furthermore, alterations in the Positive Valence Systems can be found across multiple psychiatric disorders. Besides the clear link with depression [39], impairments in reward processing are also associated with compulsive behaviors common to multiple disorders, such as obsessive-compulsive disorder (OCD), addictions, and binge eating. This has been observed with functional magnetic resonance imaging (fMRI) tasks such as the monetary incentive delay task, where patients from these populations have shown greater risk-taking tendencies and reduced loss aversion linked to abnormal activation of the striatum [47, 48]. The greater degree of reward seeking in these populations may contribute to repeating certain actions more and more frequently, which then become habits [49]. Moreover, besides alterations in reward processing in anxiety-related disorders, paralleled by alterations in frontoparietal and fronto-thalamic connectivity [50, 51], patients with OCD also show significantly higher cortisol levels than controls [52]. Literature is lacking though trying to disentangle the complex interaction between cortisol reactivity, reward processing, and compulsive behaviors, as well as their associated neural correlates.
The study by Den Ouden et al. represents an attempt to tackle this [53]. They used a transdiagnostic approach to identify compulsivity subtypes based on multidimensional markers (see Table 1), and characterized whole-brain resting-state FC. They found three subtypes with varying levels of compulsivity, behavioral avoidance, stress (including the cortisol awakening response – CAR), and learning bias, which in turn presented a different pattern of connectivity between the bilateral amygdala and the rest of the brain. This study illustrates the ability to capture neurobiological distinctiveness using transdiagnostic approaches. Moreover, it shows that those subtypes with higher stress levels are more likely to present behavioral avoidance, which highlights the relevant role of cortisol reactivity in the interplay between reward processing and compulsive behaviors.
Cognitive Systems
Cognitive processes are central to the experience of anxiety, with impairments across different cognitive domains linked to its severity [16, 54]. Individuals with high levels of anxiety exhibit difficulties in attention, memory, interpretation, and expectancy processes, biased toward anxiogenic information [55]. This contributes to increased engagement with emotionally salient stimuli, difficulties in disengaging from them, and disruptions in overall information processing [56].
For instance, anxiety severity in clinical and non-clinical samples is related to higher attentional fixation on threatening stimuli, which been linked to increased activity in emotion-related brain regions (i.e., amygdala and insula) as well as components of the ventral attention network, including the inferior parietal lobe [57]. Besides, reduced cognitive flexibility in anxiety patients has been associated with hypoactivity in brain regions responsible for top-down regulation, particularly the ACC and the dlPFC [54, 57]. This may underlie a bias toward negative stimuli both in people with high and clinical anxiety, contributing to difficulties with other cognitive control processes like goal selection, response inhibition, and adaptive behavior in emotional contexts. In this regard, cortisol is of high interest as it modulates neural activity via membrane-bound receptors in these regions: elevated cortisol levels seem to impair PFC function, in turn reducing top-down attentional control. At the same time, cortisol enhances activity in limbic and subcortical regions, especially the amygdala, thus promoting bottom-up, stimulus-driven attention through the salience network. This is a way for prolonged stress to further weaken PFC regulation of the amygdala, reinforcing a shift from goal-directed cognitive control to reactive, salience-driven processing [13].
Anxiety is also related to memory-related difficulties, which contributes to the emergence of distressing thoughts and excessive worry [58]. Emotional experiences shape memory processes, which depend on the integrity of structures rich in glucocorticoid receptors like the hippocampus and the amygdala [59]. The effects of cortisol on memory have been documented in both healthy individuals and those diagnosed with anxiety disorders. Elevated cortisol interacts with hippocampal-dependent systems, negatively influencing hippocampal volume and memory performance [60, 61] while enhancing the encoding of vivid threatening stimuli, contributing to the vivid and persistent anxiety-related memories [62] (Table 1).
Another critical anxiety-related process is cognitive control. Difficulties in this process are associated with the sense of uncontrollability during anxiogenic situations and the impaired inhibition of “automatic” responses to perceived threats or repetitive rumination. In line with this, Zhao et al. examine the association between high levels of trait anxiety and cognitive emotion regulation [63]. They found that individuals with high trait anxiety show reduced activity in the cognitive control network, including the dlPFC, ACC, and posterior parietal cortex, which aligns with prior findings on impaired top-down emotional regulation. Other processes, like goal-directed behavior, are influenced by both others’ and individual emotional states, which can trigger automatic responses such as approach or avoidance behaviors [64]. Lapate and colleagues emphasize the role of the lateral frontal pole (lFP) as a critical hub where emotional context shapes the representation of action goals, together with other prefrontal regions like the dlPFC. These conjunctive representations enable behavioral flexibility to changing environmental demands. In individuals with high anxiety, the amygdala-lFP pathway appears to be hyperactive, leading to increased excitability and a disruption in the excitation/inhibition balance within the lFP during emotional situations. This imbalance may impair the capacity to flexibly adjust behavior based on emotional cues, fostering a dominance of amygdala-driven processing [65], which could support the bias toward emotional information found in highly anxious individuals. In summary, elevated cortisol is linked to reduced cognitive control, heightened amygdala activity, and decreased prefrontal function [66].
Social Processes
Social Processes mediate the responses to multiple types of interpersonal settings, including the perception and interpretation of your own and other’s actions. As such, global social functioning deficits, and dysfunctions in the perception and understanding of the self in specific, have been strongly linked with multiple conditions, including anxiety [16] and autism spectrum disorder [67].
Higher trait-dependent baseline oxytocin levels have been associated with increased social skills, adaptive behaviours and receptive language abilities [68, 69], acting as a protective mechanism by dampening HPA and sympathetic stress activity [68] (Table 1). Children with autism spectrum disorders, however, display decreased trait-dependent oxytocin levels, generating a dysfunction of the oxytocinergic HPA-stress attenuation mechanism [68]. In turn, chronic stress induced by the continuous HPA-sympathetic hyperarousal [69, 70] results in altered FC between the PFC and the amygdala, leading to emotional dysregulation and heightened stress responses to social stimuli [71], similarly to what can be observed in OCD patients [72]. Therefore, this functional dysconnectivity pattern might be contributing to the enhanced HPA axis responsivity during benign social interactions with peers and socially-charged unfamiliar situations in autistic children [69, 73]. In contrast, when exposed to social evaluative threats, children with autism spectrum disorders exhibit blunted physiological and cortisol responses with faster return to basal cortisol levels than typically developing children [69,


Comments (0)