
Remember me
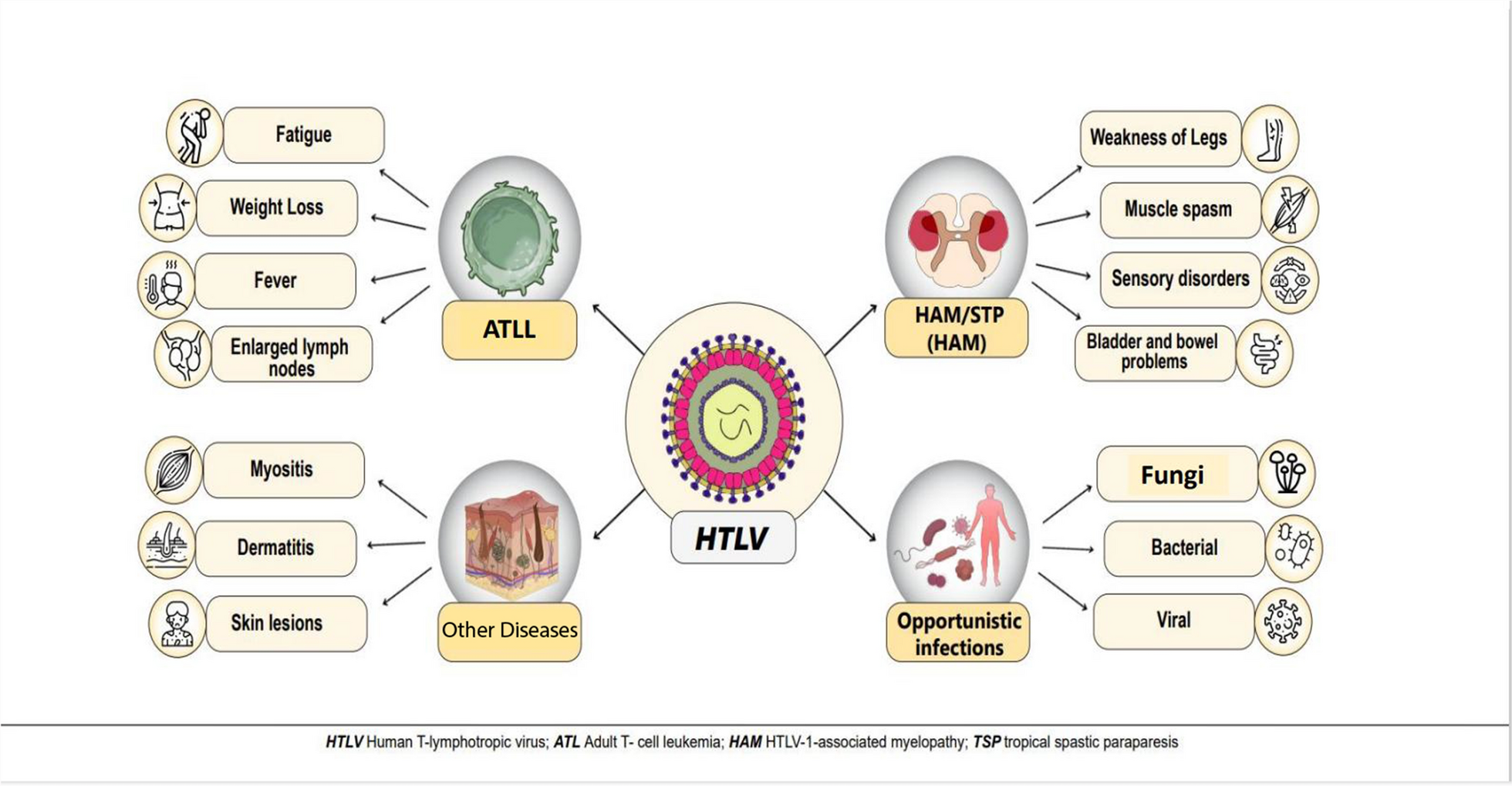

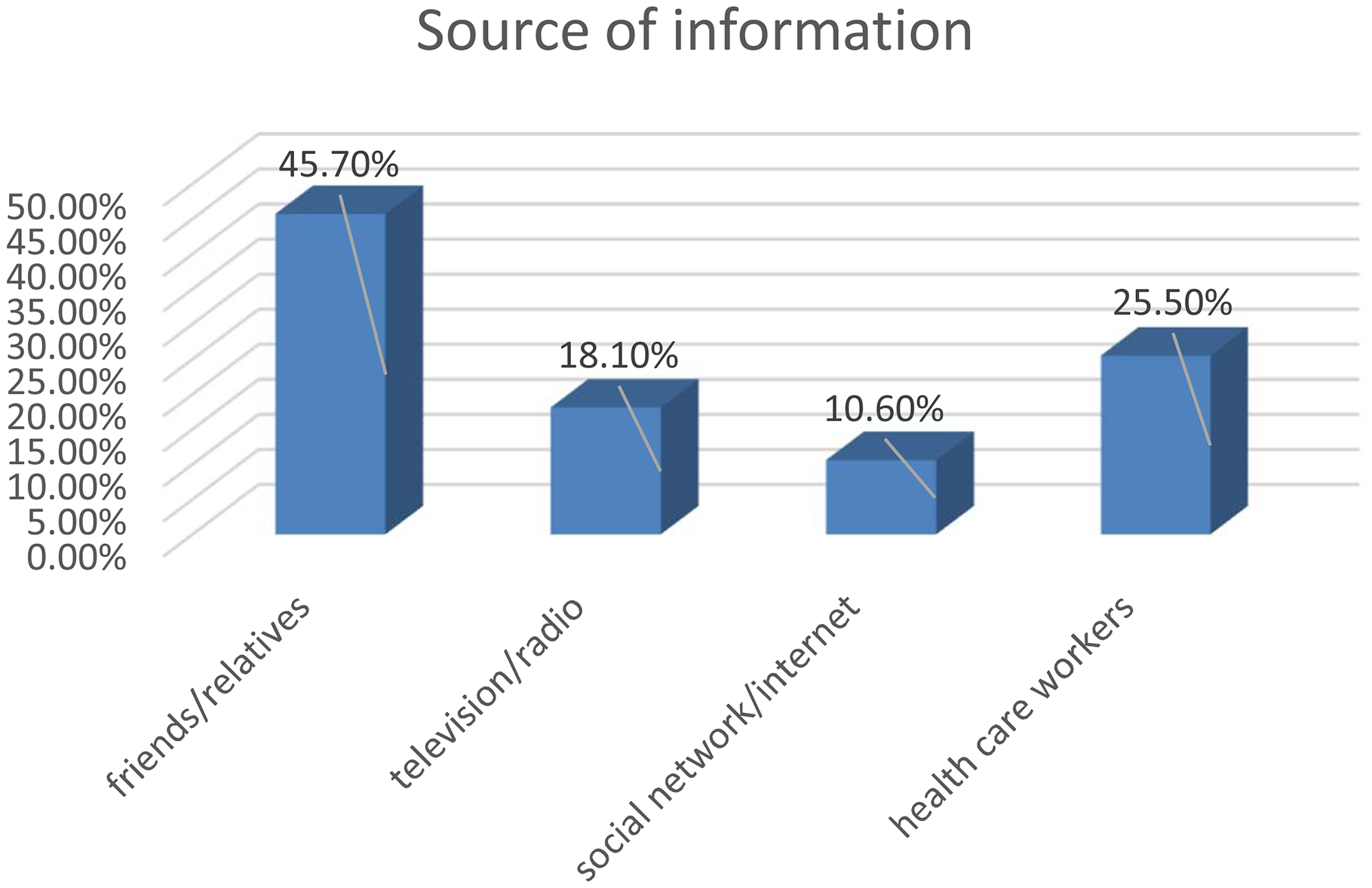


HTLV-1 is a complex retrovirus that affects over 10 million people worldwide, with endemic regions including Japan, parts of Africa, South America, and the Middle East [1]. HTLV-1 is the etiological agent of two major diseases: adult T-cell leukemia/lymphoma (ATLL or ATL) and HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) [2]. ATLL is an aggressive form of T-cell malignancy, while HAM/TSP is a chronic neuroinflammatory disorder marked by progressive spastic paraparesis and other neurological impairments [3,4,5]. Despite significant advancements in understanding HTLV-1 pathogenesis, the intricate mechanisms underlying disease progression and persistent infection remain incompletely understood. MicroRNAs (miRNAs) have emerged as key regulators of post-transcriptional gene expression, impacting various physiological and pathological processes [6]. Dysregulation of miRNA expression has been implicated in numerous diseases, including cancer, viral infections, and autoimmune disorders. Recent studies highlight the involvement of miRNAs in the complex interactions between HTLV-1 and its host, revealing novel mechanisms underlying HTLV-1 pathogenesis [7, 8] (Fig. 1).
Fig. 1
This figure briefly shows HTLV-1 and related diseases. The main diseases of the virus are HAM/TSP and ATLL, and also connections of the virus with other diseases such as a variety of skin and muscle diseases have been found
HTLV-1 encodes key regulatory proteins including transactivator protein (Tax) and HTLV-1 basic leucine zipper (HBZ), which play crucial roles in viral replication and cellular transformation. These viral proteins modulate host cellular pathways and miRNA expression profiles, creating an environment favorable for viral persistence and disease progression [9].
HTLV-1 infection also induces dynamic changes in host miRNA expression, affecting critical pathways related to immune response modulation, oncogenesis, and neuroinflammation. Understanding these interactions is essential for developing targeted therapies against HTLV-1-associated diseases [7]. MicroRNAs significantly contribute to the oncogenic transformation associated with HTLV-1 infection [7]. miR-155 and miR-21 have oncogenic properties by promoting cell proliferation and inhibiting apoptosis [10]. Moreover, when the let-7 family is downregulated, it can no longer suppress oncogenes like HMGA2, which in turn promotes cellular transformation [11, 12] (Fig. 2). miR-223 and miR-150 also modulate key oncogenic pathways, including STAT1 regulation, as shown by Moles et al. The dysregulation of these miRNAs creates a tumor-promoting microenvironment, highlighting their role as potential therapeutic targets in ATLL [13].
Fig. 2
The relationship between two key viral genes, Tax and HBZ, and dysregulation in miRNAs, as well as the relationship between some MiRs and immune system disorders, is shown in this figure
Emerging evidence suggests that miRNAs may influence early events in HTLV-1 infection, including viral transmission and entry into target cells. Though direct studies are limited, miRNAs have been implicated in the regulation of receptor expression, such as GLUT1 and NRP-1, which mediate HTLV-1 entry [14,15,16,17]. Furthermore, some miRNAs may affect synapse formation and cell-to-cell viral transfer, facilitating efficient HTLV-1 transmission and persistence. These findings highlight a potential, yet underexplored, role of miRNAs in HTLV-1 spread [17, 18].
The aim of this article is to offer a thorough perspective on the present understanding and potential developments concerning miRNAs' involvement in HTLV-1 transmission, fostering further research in this field and opening new avenues for the management of HTLV-1-related pathologies.
Dysregulated miRNAs in HTLV-1 infectionThe virus utilizes various mechanisms or tactics to establish persistent infection and evade host immune surveillance. The viral regulatory protein Tax occupies a central position in viral replication and cellular transformation [19]. HTLV-1 employs its regulatory proteins to manipulate host cellular processes, particularly through the modulation of miRNA expression. The viral protein Tax activates some viral gene expression and significantly influences host pathways, including those governing immune responses and cell cycle control. Research indicates that Tax can alter miRNA expression profiles, leading to the disruption of specific miRNAs involved in cell survival and immune regulation [20]. Additionally, HBZ protein has been implicated in the dysregulation of miRNA-mediated gene regulation. This dysregulation promotes T-cell proliferation and survival, contributing to the pathogenesis of HTLV-1-associated diseases [21]. These findings underscore the critical roles of Tax and HBZ in modulating host miRNA networks, thereby facilitating viral persistence and disease progression (Fig. 2).
Studies have identified specific miRNAs dysregulated in HTLV-1-infected cells. One of the well-characterized miRNAs is miR-155, which is upregulated in HTLV-1-infected individuals and targets critical immune regulatory molecules, including SOCS1, IL-6, PGE2, TSG6 and bcl-2 [22,23,24,25] (Fig. 2). The dysregulated miRNA, miR-21, promotes cell proliferation and resistance to apoptosis by attacking genes that inhibit tumor growth such as the cell death regulator PDCD4 [26]. These research showed that miR-21 plays critical role in cancer development and regarding HTLV-1, we can also focus on miR-21. The let-7 family of microRNAs has also been implicated in HTLV-1 pathogenesis. Let-7 miRNAs are frequently downregulated in HTLV-1-infected cells and ATLL [27]. Let-7 targets oncogenes, including high mobility group AT-hook 2 (HMGA2) and Lin-28 homolog A (LIN28A), which are involved in cellular transformation and proliferation [28, 29] (Fig. 2).
In addition to previously discussed miRNAs, Nascimento et al. conducted a pilot study demonstrating a global dysregulation of non-coding RNAs, including small RNAs, in ATLL patients. Their findings underscore the significance of miRNA profiling in HTLV-1 pathogenesis [30]. Another study by Bellon and Nicot highlighted the overexpression of H19/miR-675 in ATLL and its association with Notch signaling, further supporting the oncogenic role of miRNAs [31]. Fayyad-Kazan et al. identified circulating miRNAs such as miR-29c and miR-885-5p as potential non-invasive biomarkers for HTLV-1 diagnosis [32].
MicroRNAs and immune dysregulation in HTLV-1 infectionHTLV-1 infection causes dysregulation of immune response pathways, which are crucial for viral clearance and infection control [33]. miRNAs have a significant role in immune responses modulation during HTLV-1 infection [34]. miR-155 targets SOCS1, a negative regulator of cytokine signaling, leading to enhanced T-cell proliferation and activation [35]. miR-146a, which is upregulated in HTLV-1-infected cells, targets key molecules involved in Toll-like receptor signaling, resulting in the dysregulation of innate immune responses [36, 37].The dysregulated miRNAs support to the enduring presence of HTLV-1 infection by evading immune surveillance and creating an immunosuppressive microenvironment [20]. Moreover, HTLV-1-infected cells exhibit decreased levels of miR-150, which plays a role in regulating the activation and differentiation of T-cells. This may contribute to the assertion that immune dysregulation occurs in HTLV-1 infection [38].
Studies have indicated that miR-27a plays a role in regulating cell survival and proliferation in various cancers and viral infections. While direct evidence linking miR-27a to HTLV-1 infection remains limited, its involvement in similar viral infections, such as HIV-1, suggests a potential regulatory role. One of its known targets is prohibitin 1 (PHB1), a tumor suppressor protein involved in cellular senescence and apoptosis [39, 40].
miR-223 plays a crucial role in immune modulation and cellular transformation. It regulates immune cell differentiation by targeting genes like NFIA, C/EBPα, and Mef2c, influencing granulocyte and macrophage function. Additionally, it modulates inflammation through the NF-κB pathway, preventing excessive immune responses [41]. In the context of HTLV-1 infection, miR-223 has been shown to function as a tumor suppressor, inhibiting the proliferation of transformed T-cells, thereby reducing viral persistence and oncogenic transformation[38].
Furthermore, miR-34a and miR-146a have also been implicated in HTLV-1 infection, modulating immune responses and cellular transformation [36, 42].
Inflammatory responses and neuro-inflammationHAM/TSP is characterized by chronic inflammation and progressive spastic paraparesis [4, 43]. miR-223, which is downregulated in HTLV-1-infected cells, contributes to neuroinflammation by targeting important molecules involved in neuronal function and inflammation regulation [44, 45]. Furthermore, miR-34a has been associated with neuronal apoptosis and neurodegeneration [46, 47].
In addition, miR-155 and miR-21 promote inflammation by targeting negative regulators of inflammation, such as SHIP-1 and PDCD4, respectively. The dysregulated miRNAs contribute to the chronic inflammation seen in infected people [13, 48, 49].
Recent studies have identified other miRNAs involved in HTLV-1-associated neuroinflammation. For instance, Nicolete et al. reported upregulation of miR-125b in both asymptomatic carriers and HAM/TSP patients, suggesting a potential biomarker for disease progression [50]. Bekenstein and Soreq described the role of hnRNP A1 in neurodegenerative processes, regulated in part by miRNAs, including miR-125 and miR-223 [51]. Furthermore, de Souza et al. identified miRNAs such as miR-21 and miR-146a as potential prognostic markers in HAM/TSP patients. These miRNAs play crucial roles in controlling inflammatory cascades and neuronal survival [52].
Comments (0)