
Remember me
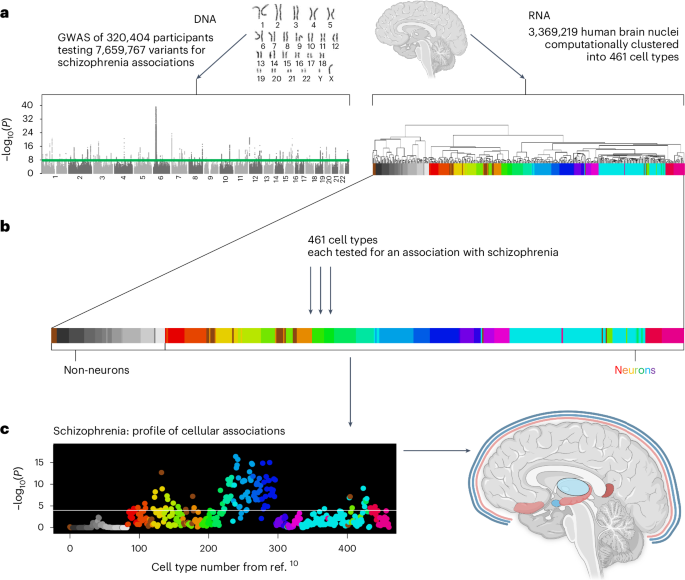


Of the 109 cell types that were significantly associated with schizophrenia, there were ten relatively independent, significant cell types (Fig. 2a). The most significant was a subtype of somatostatin (SST) interneurons (no. 239, P = 4.3 × 10−17). The next two were also cortical: PAX6 interneurons distributed widely across the cortex (no. 278, P = 1.5 × 10−15) and an excitatory cell type found almost exclusively in the retrosplenial cortex (no. 132, P = 2.1 × 10−13, 91% of cells from the retrosplenial cortex); no. 132 is probably from cortical layer 5, as per ref. 10. Note that the PAX6 interneurons (no. 278) were annotated as GABA/VGLUT3, indicating coexpression of GABA and glutamate, which is relatively uncommon. Fourth and fifth were two distinct inhibitory amygdala neuron types (no. 233, P = 2.8 × 10−12 and no. 423 (P = 9.0 × 10−10)). The remaining five cell types had neurons primarily from the prefrontal cortex (no. 404, P = 7.3 × 10−8; Brodmann area 14), thalamus (no. 440, P = 1.4 × 10−5), cortex-wide excitatory neurons annotated to deep layer 6b (no. 98, P = 2.2 × 10−5) and two excitatory hippocampal neuron types (no. 179 P = 2.6 × 10−5 and no. 202 P = 1.1 × 10−4). In addition to these ten relatively independent, significant cell types, other notable significant cell types were medium spiny neurons in the striatum (that is, caudate and putamen, no. 222), cortex-wide excitatory neurons in layer 2/3 (no. 123), and cell types preferentially located in the visual cortex (no. 133), septal nuclei (no. 428), superior colliculus (nos. 433 and 367) and substantia innominata (no. 232). For the full results, see Supplementary Table 1.
Fig. 2: Cell type associations for schizophrenia.
a, Schizophrenia results for 461 cell types are depicted in a scatterplot, with cell type number on the x axis and statistical significance on the y axis (−log10(P)), such that higher values are more statistically significant. The horizontal gray line denotes Bonferroni correction for the 461 regression analyses (P < 0.0001). The numbers above the points are cell type numbers; the numbers within the points are supercluster numbers. Larger points denote the ten cell types that were identified as relatively independent using conditional analyses. b, The top cell type for schizophrenia (no. 239) aligned to the SST interneuron subtype SST.ix (from ref. 24) for which cortical layer localization was determined using spatial transcriptomics. Thus, we inferred that cell type no. 239 was probably most abundant in cortical layer 5, but also found in cortical layers 6 and 2 and 3, whereas cell type no. 242 was probably most abundant in layers 2 and 3. Panel b adapted with permission from ref. 23, AAAS.
Interneurons associated with schizophrenia match and extend previous findingsFor schizophrenia, arguably the most widely replicated postmortem tissue finding involves aberrant inhibitory neurons in the cortex (interneurons)22,23. While both the parvalbumin and SST subclasses of interneurons have been implicated, the most recent evidence showed the greatest abnormalities in SST interneurons22,23. Thus, the present report of the strongest association with SST interneurons, and many significantly associated interneurons (including parvalbumin), is concordant with leading previous findings. We extended these findings by specifying more subtle subtypes of SST interneurons and by determining the probable cortical layers for these schizophrenia-associated SST interneuron subtypes (nos. 239 and 242) using additional snRNA-seq and spatial transcriptomics datasets. We also examined cell type no. 242 because it was nearly as significant as the top cell type (no. 239). This second-most significant cell type (no. 242) was also associated with other psychiatric phenotypes, included ~3× more cells than no. 239 and it was the most significant cell type in our analysis of the 2018 schizophrenia GWAS. Thus, we thought no. 242 was also a strong candidate for psychiatric relevance.
Recent studies identified far more interneuron subtypes than were distinguishable using traditional markers. These transcriptomically defined cell types reside preferentially in specific cortical layers in both humans and mice24. Figure 2b, using a plot adapted from ref. 24, shows the cortical layer distribution of human interneuron subtypes (that is, cortical layers 1–6). We aligned SST subtypes and found that nos. 239 and 242 (from ref. 10) best matched SST.ix and SST.i, respectively (from ref. 24), and that these cell types differed in cortical layer distribution. No. 239 SST interneurons were most likely to be localized to cortical layer 5 (less so to layers 6 and 2/3), while no. 242 SST interneurons were most likely to be in layers 2/3. Alignment to two mouse datasets25,26 yielded similar cortical layer localization results, providing additional support for putative layer localizations. Contextualizing these findings, we note that schizophrenia postmortem tissue studies have primarily identified deficits in upper cortical layers, particularly deep layer 3 (ref. 27), which may be consistent with the primary localization of cell type no. 242 and, to a lesser extent, no. 239. Given known disruptions in sensory processing and integration in schizophrenia, it is conceivable that cortical layer 5 integration and output functions28 are disrupted in schizophrenia because of abnormalities in layer 5 SST interneurons of the no. 239 type.
Subcortical neuronal associations with schizophreniaIn the amygdala, a structure known to have diminished volume in schizophrenia29, we found 17 significant cell types (including both inhibitory and excitatory), two of which were independent significant cell types (nos. 233 and 423, both inhibitory). Cell type no. 233 was annotated by Siletti et al.10 as a medium spiny neuron of the eccentric subtype, a recently discovered subtype of medium spiny neurons9. This general class of neurons—medium spiny neurons—has been frequently linked to schizophrenia; medium spiny neurons are the dominant inhibitory neuron of the striatum (caudate and putamen). In addition to the two main subtypes of medium spiny neurons, which are well characterized based on their differential expression of dopamine receptors (D1 versus D2), recent transcriptomic studies made it clear that a third subtype of medium spiny neurons also exists. These ‘eccentric’ medium spiny neurons evaded detection because classical D1 and D2 markers do not reliably differentiate the newly named ‘eccentric’ medium spiny neurons9. Thus, linking this eccentric medium spiny neuron type (no. 233) to schizophrenia demonstrates the multiple benefits of large-scale transcriptomic studies and the unbiased approaches used in this study to link cell types to schizophrenia. First, scRNA-seq studies made possible the detection of the relatively rare (~4%) but clearly transcriptomically distinct eccentric medium spiny neuron subtype9. Second, the brain-wide snRNA-seq dataset from ref. 10 used in this study shows that neurons that transcriptomically resemble all three subtypes of medium spiny neurons (D1, D2 and eccentric) are also found outside the striatum. Third, this approach links these newly discovered, relatively rare and extrastriatal eccentric medium spiny neurons to schizophrenia. Such a discovery would not have been possible without these large, comprehensive and unbiased datasets that allow us to see more clearly the gaps in previous knowledge and to discover which newly filled gaps might also provide critical information about schizophrenia etiology. Consistent with previous research, we found striatal associations of medium spiny neurons (nos. 222 and 224).
The second relatively independent, significant amygdala cell type (no. 423) is inhibitory; while found predominately in the amygdala (51% of cells), it is also present in the thalamus (20%), hypothalamus (15%) and other brain regions. Other amygdala cell types associated with schizophrenia, but not deemed independent after conditional analyses, include the medium spiny neurons of the D1 (for example, no. 220) and D2 (no. 217) subtypes.
Our hippocampal findings provide molecular details that can augment and extend schizophrenia imaging and postmortem tissue findings. In this study, we identified specific neuron types that may underlie these hippocampal volume reductions in schizophrenia. Of the 86 neuron types located primarily in the hippocampus (using data from ref. 10), seven were significantly associated with schizophrenia in our analyses. Of these, conditional analyses found two relatively independent cell types (nos. 179 and 202). Cell type no. 179 is an excitatory neuron type (VGLUT1); contributing cells came from all major subregions of the hippocampus. Cell type no. 202 is also excitatory (VGLUT1); contributing dissections include all major hippocampal subregions except the subiculum. The present findings may be concordant with previous reports of increased hippocampal excitatory (glutamate) neurotransmitter metabolites in individuals with schizophrenia30.
In summary, the primary anatomical locations for the relatively independent, significant schizophrenia cell types were three widely distributed across the cortex, two from specific cortical regions (retrosplenial and prefrontal), two from the amygdala, two from the hippocampus and one from the thalamus. The three subcortical structures linked to schizophrenia in this study (amygdala, hippocampus and thalamus) are precisely the same subcortical structures with the largest volume decreases in patients with schizophrenia compared to controls (Hedges g of −0.66, −0.46 and −0.31, respectively) according to a meta-analysis of brain volume studies in first-episode psychosis29. Thus, two independent, brain-wide, data-driven approaches to understanding schizophrenia—one using imaging data29 and the present approach using genetic data—pointed to the same three subcortical regions: amygdala, hippocampus and thalamus. In the next section, we consider the cell types and brain regions linked to four other phenotypes.
Contrasting cellular associations for five brain phenotypesWe sought to establish whether this approach could accurately link cell types to informative complex brain phenotypes: alcohol consumed per week; sleep duration per night; multiple sclerosis; and Alzheimer’s disease. These phenotypes were selected for the presence of expected cellular associations and for the existence of adequately powered GWAS for these phenotypes. Figure 3 shows that the expected cell types were significantly associated with these phenotypes (see Supplementary Tables 2–5 for the full cell type results, Extended Data Figs. 1–4 for the larger annotated scatterplots that compare the phenotypes and Supplementary Tables 6–10 for the gene-level results).
Fig. 3: Distinct cell type profiles for five phenotypes.
Cell-type associations for five brain-related phenotypes depicted in scatterplots, with cell types depicted in order (for 461 cell types) along the x axis and with statistical significance on the y axis (reported as −log10(P), such that higher values are more significant). The horizontal white lines represent the Bonferroni correction for 461 regression analyses (that is, P < 0.0001). The first three phenotypes (schizophrenia, alcohol consumed per week and sleep duration per night) yielded neuronal cell type associations. The two neurological phenotypes (multiple sclerosis and Alzheimer’s disease) yielded nonneuronal cell type associations of immune and microglial cell types, respectively. See Extended Data Figs. 1–4 for larger annotated scatterplots that compare the phenotypes.
For alcohol consumed per week, the top cell type was a D2 medium spiny neuron (no. 217, P = 1.3 × 10−9), which matched previous expectations given that D2 medium spiny neurons causally influence alcohol consumption31,32,33,34,35. For sleep duration per night, the top cell type was also a medium spiny neuron, but of the D1 type (no. 231, P = 2.8 × 10−9). This finding may be consistent with recent findings linking D1 medium spiny neurons and sleep, particularly rapid eye movement sleep36. For sleep duration per night, we also highlight associated cell types from the pons (no. 396, P = 1.3 × 10−6) and medulla (no. 386, P = 5.9 × 10−6) as these structures are key nodes of sleep regulatory circuits. Note that these pons and medulla cell types were not associated with schizophrenia, alcohol consumption nor any other psychiatric phenotypes tested; rather, they are specific among these phenotypes for sleep. The sleep phenotype had many associations with the newly named Splatter supercluster of neurons (described in ref. 10), which includes a wide variety of subcortical neuron types (see Fig. 4 for a comparison of the brain regions harboring the relatively independent, significant cell types for these three phenotypes).
Fig. 4: Brain locales of origin for the relatively independent significant cell types associated with schizophrenia, alcohol consumed per week and sleep duration per night.
Inhibitory cell types are depicted in blue, excitatory cell types are depicted in red, with color saturation denoting statistical significance from the regression analyses. Unless an asterisk is present, the regions depicted were the source of more than 50% (and usually much more than 50%) of cells for a particular cell type. If there are more than one excitatory or inhibitory cell types for a single brain structure, color saturation corresponds to the more significant cell type. If both excitatory and inhibitory cell types are associated with a single structure, then the fill of the structure (as opposed to the border) denotes the more significant association. For example, there are two amygdala cell types associated with alcohol per week and the inhibitory cell type no. 217 is more significant (hence the blue/inhibitory fill). Cell type no. 278 is annotated as GABA/VGLUT3. VIP, vasoactive intestinal peptide. Figure created with BioRender.com.
For multiple sclerosis, an autoimmune condition, the strongest association was a T cell type (no. 1, P = 6.0 × 10−20). The next most significant cell types were B cells (no. 0, P = 4.7 × 10−14) and natural killer cells (no. 2, P = 3.5 × 10−12). These results are consistent with longstanding understanding of pathogenic T cell involvement in multiple sclerosis, the best-available treatments for multiple sclerosis and also more recent findings linking B cells to the disease37,38,39. For Alzheimer’s disease, the most significant cell types were microglial (most associated cell type: no. 6, P = 2.4 × 10−7). Previous work established that many Alzheimer’s disease genes are preferentially expressed in microglia; newer therapeutics targeting microglia are being investigated40,41.
Data and analytical requirements for sufficient statistical powerWe sought to determine how the statistical power of individual GWAS influenced our detection of cell type associations, hypothesizing that a poorly powered GWAS would not afford discovery of cellular associations. We further hypothesized that successive GWAS with increasing statistical power (as evidenced by the increasing numbers of loci detected) would afford increasing numbers of associated cell types until most of the relevant cell types were associated, at which time the results would appear to plateau (that is, diminishing discovery of cell types as sample sizes continued to increase). Thus, we reran our primary analysis on four schizophrenia GWAS with progressively increasing sample sizes (and statistical power), ranging from a 2011 GWAS (n = 51,695, effective n = 46,729) to the most recent GWAS (n = 320,404, effective n = 233,471)6,42,43,44. Figure 5 shows results that are consistent with our hypotheses. The smallest GWAS revealed no significant cell type associations in our analysis, although it was adequately powered to detect seven GWAS loci. By contrast, the subsequent schizophrenia GWAS afforded detection of 63, 94 and then 109 cell types, corresponding to the discovery of 108, 145 and then 287 loci as reported in primary publications6,42,43. Our results suggest that plateauing of cell type discovery may occur at much smaller sample sizes than required for GWAS loci (predicted to saturate at more than 10,000 associated variants for psychiatric disorders). This also suggests that relevant biological features are being captured at the cell-type-level, via aggregation of polygenic signal into cell types.
Fig. 5: Required statistical power of a GWAS for detecting cell type associations.
a, Number of cells in each cell type. b, Results for four successive, increasingly large and better powered schizophrenia GWAS. The horizontal lines in all plots denote the Bonferroni-corrected statistical significance for 461 regression analyses (P < 0.0001). c, In this range of sample sizes for schizophrenia GWAS, the number of loci detected continued to increase steeply, but the number of cell types plateaued. Effective n accounts for the decrease in statistical power attributable to imbalanced case control ratios: neff = 4/((1/cases) + (1/controls)). IQR, interquartile range.
The statistical power for these analyses also depends on the accuracy of gene expression measured in individual cell types. Cell types with lower numbers of cells tend to have less accurate values for average gene expression. Thus, we first tested whether statistical significance was associated with the number of cells, and it was not (r = 0.06, P = 0.18 for the correlation between schizophrenia (−log10(P)) and the number of cells) (see Extended Data Fig. 5a for the graphical depiction). Considering the wide range of cells per cell type (minimum = 34, maximum = 124,837), we also progressively downsampled to assess the effect of lower numbers of cells, in individual cell types, on the power to detect associations with schizophrenia (that is, random sampling of cells in each cell type). Based on the minimum number of cells in any cell type (ncells = 34), we downsampled three times to 3,400, 340 and 34 cells (these are maximum values given that some cell types did not have 340 or 3,400 cells). Extended Data Fig. 5b shows that the pattern of associated results was relatively stable across downsampling analyses. Indeed, 56 of the original 109 cell types were significant even when downsampling to just 34 cells per cell type.
We also sought to understand the influence of method-dependent factors on our results. The LDSC-SEG21 results are given in Supplementary Table 23. We observed reasonably high consistency between our primary analysis method (MAGMA gene property analysis) and LDSC-SEG: Spearman’s rank correlation for schizophrenia (r = 0.71), alcohol per week (r = 0.62) and sleep duration per night (r = 0.72). However, differences also existed. LDSC-SEG identified no significant cell types for MS despite the established involvement of immune cells; similarly no cell types were associated with AD. The LDSC-SEG analysis also failed to identify D2 medium spiny neurons for alcohol consumption and the pons and medulla cell types for sleep duration per night. The results suggest that LDSC-SEG may not be as well powered as the MAGMA gene property analysis.
Progress toward a data-driven cellular taxonomy for psychiatric disordersThis work suggests the possibility of a taxonomy for psychiatric disorders based on quantitative evaluation of cell types (Fig. 6a). A cellular taxonomy could be valuable for structuring future exploration; for example, we can infer biological characteristics of each cell type from transcriptomic information. Figure 6b shows receptor and neuropeptide usage as inferred from transcriptomic data. Further, Extended Data Fig. 6 depicts an in silico exploration of predicted drug-cell type pairings, meaning cell types that putatively are influenced by specific drugs. We also sought to characterize the biological processes, cellular components, and molecular functions of individual cell types via overrepresentation analysis of Gene Ontology (GO) categories (see Methods for details and Supplementary Tables 18–22 for results).
Fig. 6: A cellular taxonomy of brain phenotypes maps shared and non-shared cellular associations in a mechanistically informative manner.
The 31 superclusters are color-coded as in Fig. 2. a, Relatively independent significant cell types are depicted for the five primary phenotypes plus bipolar disorder and depression. Shared or ‘pleiotropic’ cell types include cortical interneurons (no. 239) and excitatory retrosplenial cortex neurons (no. 132). Regarding non-shared cell types, sleep-specific associations of pons (no. 396) and medulla (no. 386) splatter neurons are notable. b, Experimental follow-up may include testing of drugs as predicted by gene expression in associated neuron types. 1Selected receptor genes among the 200 genes with highest specificity values, for a given cell type. 2Selected neuropeptide autoannotation results from ref. 10.
Extension to other psychiatric phenotypesRegarding other psychiatric disorders, bipolar disorder and depression cell types overlapped, but not entirely, with schizophrenia cell types (Fig. 6, Supplementary Tables 11 and 12 and Extended Data Figs. 7–9). Regarding interpretation, it is important to note that published schizophrenia GWAS identified more significant loci and afforded detection of more cell types (109 cell types) than any other psychiatric disorder in this study, including bipolar disorder (83 cell types) and depression (41 cell types). Further, genetic correlations and polygenic scores suggest that phenotypic heterogeneity is probably higher in bipolar disorder and depression datasets than with schizophrenia. Consequently, cell types for bipolar disorder and depression may be less accurate than for schizophrenia because of a variety of factors, including statistical power, true differences in phenotypic heterogeneity and differences in phenotyping quality across phenotypes, thus requiring further study. For other psychiatric disorders, only a few cell types (or no cell types) exceeded statistical significance (P < 0.0001). This is likely because of insufficient power in the currently available GWAS for anorexia (no cell types), substance use (no cell types), PTSD (no cell types), autism (no. 428) and ADHD (cell types nos. 136, 234, 377 and 228), and probably not because of a lack of truly associated cell types (for the full results, see Supplementary Tables 13–17).
Comments (0)