
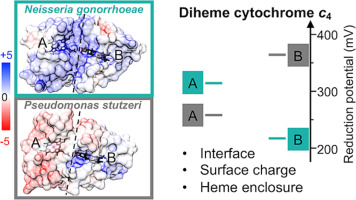
Remember me

In photosynthetic organisms, the process of electron transfer is fundamental for converting solar energy into electrochemical and chemical energy. This process requires coordinating multi-component reactions across extensive spatiotemporal networks, resulting in a robust and complex solar conversion system that powers energy conversion reactions. Iron‑sulfur (FeS) proteins have an essential role in numerous biological processes and can function as catalysts or electron carriers to support cellular metabolism including photosynthesis, respiration, nitrogen fixation, and stress responses, along with other functions [[1], [2], [3]]. Ferredoxins, small molecular weight FeS proteins found ubiquitously across species, play a crucial role as electron carriers in energy conversion networks.
The cyanobacterium Synechocystis sp. PCC 6803 (S. 6803) serves as a valuable model organism for studying the biochemical components and networks that couple photosynthetic water-oxidation chemistry to reduction-oxidation reactions. The S. 6803 genome encodes nine proteins with characteristic FeS-binding motifs, of which Fdx1 (i.e., PetF, ssl0020) is the most abundant [[4], [5], [6]]. The expression levels of the other eight Fdxs vary depending on environmental conditions, such as fluctuating light, CO2 concentration, or redox and oxidative stress [4,7]. Some of the better characterized Fdxs include Fdx1, Fdx2 (sll1382), Fdx4 (slr0150) [8], [[9], [10], [11], [12], [13]] and Fdx7 (sll0662) [14,15]. However, there are still substantial gaps in our understanding of their structural and functional characteristics, as well as their roles in biological processes and interactions with protein partners. Most of the Fdxs in S. 6803 are binding a single [2Fe 2S] cluster, with the exceptions being Fdx7, Fdx8 (ssr3184), and Fdx9 (slr2059), which are predicted to bind [4Fe-4S], [7Fe8S] and [8Fe-8S] clusters, respectively [4]. Ferredoxins binding [4Fe4S] clusters are prevalent among anaerobic organisms and generally are more reducing and exhibit greater susceptibility to oxygen, in contrast to the [2Fe2S] cluster-binding Fdxs.
2S] cluster, with the exceptions being Fdx7, Fdx8 (ssr3184), and Fdx9 (slr2059), which are predicted to bind [4Fe-4S], [7Fe8S] and [8Fe-8S] clusters, respectively [4]. Ferredoxins binding [4Fe4S] clusters are prevalent among anaerobic organisms and generally are more reducing and exhibit greater susceptibility to oxygen, in contrast to the [2Fe2S] cluster-binding Fdxs.
The objective of this work was to investigate the putative Fdx in S. 6803, Fdx9 and determine its essential biophysical properties and function. Fdx9 is expressed at low-abundance and is non-essential under standard photoautotrophic conditions [4]. Based on genetic studies, it is proposed that Fdx9, in conjunction with Fdx7, forms part of the ferredoxin-glutaredoxin-thioredoxin crosstalk pathway, serving a protective role in response to high-light conditions and the presence of toxic metals [4,15]. Using the bacterial adenylate cyclase two-hybrid (BACTH) system, Fdx9 was found to bind in vitro ferredoxin-thioredoxin reductase (FTR) (FtrC, sll0554), flavodiiron protein 3 (Flv3, sll0550), and 3-ketoacyl-acyl carrier protein reductase (FabG, sll0330) [4]. Proteomic studies have revealed the up-regulation of Fdx9 under high-light and low-temperature conditions, alongside other proteins involved in FeS cluster synthesis, respiratory and photosynthetic electron transport, as well as another Fdx (Fdx3, slr1828) [16]. Recent studies of a S. 6803 Fdx9 deletion strain show diminished growth under photomixotrophic conditions [17].
In summary, Fdx9 is upregulated under growth conditions that lead to redox imbalance and oxidative stress, suggesting a functional role in metabolic pathways that alleviate these stresses. The biophysical and redox properties of Fdx9 and biochemical evidence in support of the proposed functions remain open questions. To address these questions, we expressed heterologously and purified Fdx9 from S. 6803 and characterized its FeS cluster properties through a combination of electron paramagnetic resonance spectroscopy (EPR) and electrochemistry. The biophysical characterization was complimented by computational analysis of the Fdx9 structure, which suggested that Fdx9 belongs to a unique and poorly characterized group of FeS-containing proteins with two distinct domains: a ligand-binding Nil-domain and a clostridial-type [18] 2×[4Fe4S] binding domain. The N-terminal domain is a member of the NIL superfamily, which is a suggested substrate-binding domain located in the C-terminal, intracellular region of the MetN subunit of the methionine ABC transporter [19]. The results of this study, along with analysis of available genomic and bioinformatic data, are summarized to propose Fdx9 involvement in the homocysteine synthetic pathways. These findings offer new insights into the roles of two domain 2×[4Fe4S] proteins in cyanobacterial metabolism and highlight promising avenues for future research in this area.
Comments (0)