In eukaryotes, DNA polymerase α-primase (pol-prim) initiates DNA synthesis during replication, generating the first ∼30 nucleotides of nascent strands.1, 2, 3, 4, 5 Pol-prim is unique among replicative polymerases in its ability to perform de novo synthesis from a single-stranded DNA template; the primers it creates are required for further synthesis by the processive polymerases ε and δ that perform the bulk of nascent strand synthesis6, 7. Pol-prim plays a particularly crucial role on the lagging strand during replication, as primers must be repeatedly synthesized due to the discontinuous nature of synthesis of this nascent strand.8 The primers generated by pol-prim are chimeric in nature, consisting of 7-10 ribonucleotides followed by ∼20 deoxyribonucleotides.9, 10 The RNA and DNA portions of this chimeric primer are generated by distinct active sites located in the primase and polymerase subunits of this tetrameric enzyme (Figure 1).11
Primase consists of the PRIM1 (also known as p48, p49, PriS, and Pri1) catalytic subunit and the PRIM2 (a.k.a. p58, PriL, and Pri2) regulatory subunit.12 PRIM2 contains PRIM2N (residues 1-245), a domain that acts as a scaffold linking the primase subunits to the pol α subunits, and the flexibly attached PRIM2C domain (residues 265-455), which is involved in regulation of both RNA and DNA synthesis.13, 14, 15, 16, 17, 18, 19, 20 Although the catalytic site for RNA synthesis is located within PRIM1, PRIM2C greatly stimulates catalytic activity and is required for both initiation of primer synthesis and regulation of the length of the primer.21, 22, 23 Polymerase α consists of the catalytic subunit POLA1 (a.k.a. p180) and the regulatory subunit POLA2 (a.k.a. p68, p70, and B subunit).24, 25 POLA1 contains a catalytic domain (POLA1cat) and a C-terminal domain (POLA1C, residues 1265-1462) that acts as a scaffold between POLA2 and PRIM2N, while also connecting to POLA1cat through a flexible linker.17, 26, 27 Architecturally, pol-prim is organized into three modules: the tetramer core (POLA2/POLA1C/PRIM2N/PRIM1; a.k.a. pol-prim platform), POLA1cat, and PRIM2C.28, 29, 30 POLA1cat and PRIM2C are tethered to the tetramer core through ∼30 and ∼20 residue flexible linkers, respectively. The modularity of pol-prim allows for dynamic coordination of RNA and DNA primer catalysis as well as interfacing with other replication factors.31, 32, 33, 34, 35, 36, 37
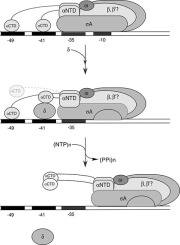
RNA primer synthesis involves the coordinated action of PRIM1 and PRIM2C. PRIM1 alone possesses an ability to elongate pre-synthesized RNA primers in the absence of PRIM2C, but is otherwise incapable of initiating primers and has low affinity for oligonucleotide substrates on its own.10, 38, 39 PRIM2C has an inherently higher affinity for substrates, in particular for structures mimicking RNA primed templates that contain a 5’ triphosphate and 3’ template overhang.28, 38, 39 It is generally understood that initiation of primer synthesis requires a configuration in which PRIM1 and PRIM2C are aligned and together bind the ssDNA template, along with two incoming NTPs and catalytic metals before synthesis of the initial dinucleotide can occur.20, 40, 41, 42, 43 Formation of the first dinucleotide is known to be the rate-limiting step of primer synthesis.10
Once the dinucleotide is formed, the RNA primer is elongated by PRIM1 in a 5’-3’ direction to a length of 7-10 nucleotides. However, PRIM1 lacks the ability to regulate primer length without PRIM2C.21, 39 Additionally, PRIM1 synthesis is distributive in nature, as synthesis of primers less than the unit length of 7 is common and is increased in the absence of PRIM2C.10, 39, 40 PRIM2C not only has higher affinity than PRIM1 for the substrate, but is also remains bound to the 5’ end of the primer, which plays an essential role in primer length regulation.39, 44, 45 From this it can be inferred that PRIM2C and PRIM1 become spatially separated as the primer length grows. Recent studies by our groups and others have also demonstrated that PRIM2C forms an interaction with POLA1cat during DNA initiation and likewise remains bound during DNA elongation, playing a critical role in handoff and DNA primer length regulation.45, 46, 47 In the prevailing model for RNA primer synthesis, a steric clash between PRIM2C and PRIM2N arises when the RNA primer reaches a length of 9 or 10 nucleotides, providing an upper limit for primer length.28, 30 Our recent work has shown that steric clash with PRIM2C prevents POLA1cat from binding an RNA primer of <7 nucleotides, providing the lower bound of unit primer length as 7.46
Despite these advances in understanding that PRIM2C regulates primer synthesis, direct structural characterization of the priming complex during RNA catalysis is lacking. Here, we report new data and present a mechanistic model for the initiation, elongation, and handoff of the RNA primer. By comparing binding affinities for RNA-primed template substrates of a variety of pol-prim constructs, we directly show that the majority of the substrate binding affinity of primase derives from the tight binding of PRIM2C. We also characterize structural transitions that occur as the RNA primer is initiated and elongated showing there is a mixture of states in all cases and that in the presence of substrate, the majority of molecules have PRIM2C but not PRIM1 bound. Our results reveal the molecular basis for the highly distributive nature of RNA primer synthesis and lead to a revised model for RNA primer synthesis that incorporates the high affinity of PRIM2C for substrate, the high-degree of flexibility of PRIM2C, and the preferential binding of RNA-primed templates by POLA1cat.


Comments (0)