
Remember me
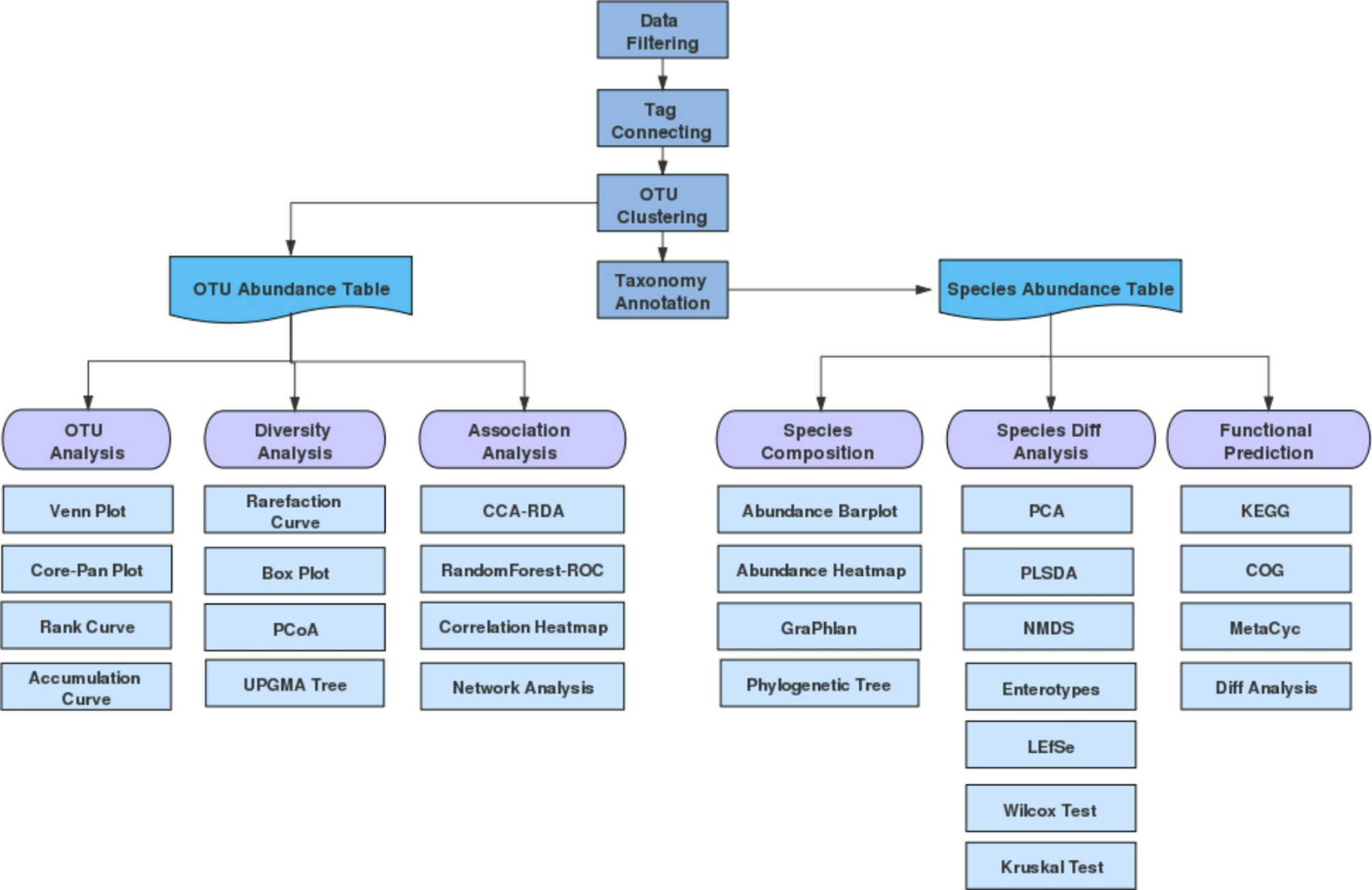






In general, StoTA was modified in two ways. We first focused on the active center, in which a lysine residue at position 282 (K282) is part of the catalytic domain that performs a nucleophilic attack on the carbonyl group of PLP to form a Schiff’s base (Crismaru et al. 2013). Under physiological conditions, the amino group of the sidechain of lysine is protonated, which makes it a weaker nucleophile. To prevent this protonation, the environment of K282 was made more alkaline. For this purpose, two amino acids in the vicinity of the catalytic lysine (I283 and I284) were replaced by glutamine. The second modification concerned the replacement of an interacting threonine residue (T317), which by forming a hydrogen bond to K282, might pull the latter away from PLP (Fig. 6). To confirm this, T317 was substituted by valine and the resulting conformational changes predicted by structural modelling (Additional file 1: Figure S2).
Fig. 6
Interaction between T317 and K282 in StoTA, data for the interactions are stored in the model 6K8H. Dotted blue lines indicate hydrogen bonds between residues and the co-factor PLP. The side chain of T317 forms a hydrogen bond to the side chain of K282, while K282 forms another hydrogen bond to the carbonyl group of PLP. Green: chain A, orange: chain B. Red dots are water molecules
In general, StoTA variants with substitutions in the active center showed a similar or lower activity as the wild type enzyme (Fig. 5). T317V was not active at all with β-homophenylalanine, β-homoalanine and β-leucine. Compared to the unmodified enzyme, the activity with β-phenylalanine also decreased to 26.3%. Only with 4-amino-pentanoic acid T317V showed a similar activity as the wild type. This confirms that residue T317 should be part of the suggested motif (Crismaru et al. 2013), or even the active center. T317 of chain A is proposed to interact with K282 of chain B, and vice versa (Fig. 6). Assuming that T317 is important for the dimerization of the enzyme, a loss of T317 will lead to a more unstable and less active protein complex. Amongst at least five other residues, T317 is also involved in PLP binding (Kwon et al. 2019). This could be another reason for the loss of activity, if the missing hydrogen bond between T317 and the phosphate group of PLP leads to an incorrect binding of the co-factor. Another possibility is that T317 is not drawing K282 away from the active center but is bending or stabilizing K282 in a position where it can bind the keto group of PLP or another substrate.
Modification of the proposed signature sequence motif in StoTAIn the second approach, StoTA was modified to match the proposed signature sequence motif that allows aminotransferases converting aromatic β-amino acids (Crismaru et al. 2013). A blast search (Altschul et al. 1997) of the putative motif sequence against the StoTA sequence revealed that only four substitutions are required to fit StoTA to the proposed motif (Fig. 3). These were S17P, T38V, Y47T and N70E. We generated fifteen different variants for this type of modification. Four of those carried a single amino acid substitution, six carried two substitutions, four carried three substitutions, and one variant carried all four substitutions. Some of the modifications of StoTA led to an increased activity, emphasizing the importance of some of the amino acids in the proposed motif. Several mutants, which were made to adapt StoTA to the motif, had comparable activities to the wild type, others showed different activity levels.
The double substitution mutant T38V|N70E led to a loss of activity, showing approximately 75% activity with 4-aminopentanoic acid and β-phenylalanine and only 35% with β-leucine, while β-homophenylalanine and β-homoalanine were deaminated at almost the same velocity as with the wild type. In contrast, the double mutant, S17P|T38V showed a strong increase in activity. Depending on the amino donor, S17P|T38V displayed a 5.4 to 9.2-fold higher activity compared to the wild type. Also, T38V|Y47T was 3.8–10.5 times more active than the wild type. A strongly enhanced activity with all tested substrates was also found in the single mutant Y47T. Depending on the amino donor, the specific activity increased by 5.0–7.0 times compared to the wild type. This might be due to the fact that threonine has a less bulky side chain than tyrosine making it easier for a substrate molecule to enter the active center.
Overall, the double mutant S17P|T38V had the highest activity of all StoTA variants with all substrates, except for β-leucine. With β-leucine the double mutant T38V|Y47T fared best with a 10.5 times enhanced activity compared to the wild type. Interestingly, as single replacement T38V had a lower activity than the wild type for all substrates except β-leucine, while S17P had lower or similar activities with all substrates. Together, however, these substitutions exerted a strong influence on the enzyme activity. This is somewhat surprising considering that T38 seems too far away from either S17 or Y47 to provoke an interaction. The increase of activity must have other reasons, a difference in protein folding can be excluded based on in silico folding.
Mutant StoTA in which N70 was replaced by glutamate had a 9.1% loss of activity with 4-aminopentanoic acid thus revealing the importance of asparagine at position 70 for transforming γ-amino acids. Regarding β-amino acids, a different influence was observed. All β-amino acids were converted faster in comparison to the wild type (β-homoalanine + 29.3%, β-leucin + 191.5%, β-phenylalanine + 94.6% and β-homophenylalanine + 33.7%). The motif proposed by (Crismaru et al. 2013) was originally suggested only for aromatic amino acids. However, as observed here, fitting StoTA to the motif had a high impact on the activity with a branched chained amino acid and a minor impact on the activity with aromatic amino acids. This shows that the motif can be used for non-aromatic ω-Tas too. N70 is located close to the phosphate group of bound PLP, where it may interact with the phosphate of the bound PLP (Fig. 7). In the vicinity of N70, R36 might be the residue, which binds the carboxyl group of the substrate to hold it in the p-pocket. Exchanging N70 by glutamate obviously alters the interactions between this residue and PLP or R36. The altered interaction appears to be beneficial for some substrates, but disadvantageous for others.
Fig. 7
Close up to the active center of 6K8H. residues of N70 and R36 are shown and named
Depending on the amino donor the quadruple mutant, S17P|T38V|Y47T|N70E, showed only 8.6–81.4% residual activity, suggesting that not all residues in the proposed motif are equally important. The highest gains of activity were achieved in the mutants Y47T, S17P|T38V and T38V|Y47T. Additionally, these mutants have a lower temperature optimum, ranging between 40 and 50 °C instead of 60 °C. This is relevant for industrial use, since a lower temperature optimum requires less energy thus reducing production costs.
Previously, the activity with β-phenylalanine was improved by site-directed mutagenesis of an ω-TA from Caulobacter crescentus (Hwang et al. 2008). Two variants of this enzyme, N285A and V227G showed a three and twofold activity increase, respectively, compared to the wild type. On the other hand, these variants had a four and twofold lower activity with β-homoalanine (Hwang et al. 2008). By introducing two mutations, (Cho et al. 2008) optimized the activity of an ω-TA from Vibrio fluvialis against aromatic and aliphatic amines. Depending on the amino donor, W57G had a 2- to 40-fold higher activity with aromatic amines and a 5- to 19-fold higher activity with aliphatic amines. In the second variant, W147G the activity with aromatic amines ranged between 0.8-fold to 14-fold relative to the wild type, while the activity with aliphatic amines ranged between 0.9- and 6-fold (Cho et al. 2008). These results are comparable to those of the best variants obtained in the present study. Although none of our variants reached a 40-fold activity gain as obtained for the V. fluvialis enzyme for one substrate (Cho et al. 2008), the best variants produced in this work showed a consistent activity rate across all substrates tested.
Specific activity with different substratesAll variants of StoTA catalyzed the deamination of all tested substrates. The activities were measured with β-homophenylalanine, β-homoalanine, 4-aminopentanoic acid, β-leucine and β-phenylalanine. The highest specific activity with β-homophenylalanine was observed with the S17P|T38V variant in which 1 µg protein was able to convert 3.211 ± 0.009 µmol β-homo-phenylalanine in 1 min, which is defined as U µg−1. Y47T has a quite similar activity, 3.007 ± 0.017 U µg−1, and T38V|Y47T was able to deaminate 2.305 ± 0.026 µmol β-homo-phenylalanine per minute.
For β-homoalanine, S17P|T38V achieved the highest specific activity, namely 2.226 ± 0.208 U µg−1, followed by Y47T, 1.981 ± 0.043 U µg−1 and T38V|Y47T with a specific activity of 1.591 ± 0.112 U µg−1. With 4-aminopentanoic acid, S17P|T38V was the only variant with a specific activity above 1 U µg−1 (1.146 ± 0.037 U mg−1). With an activity of 1.369 ± 0.080 U µg−1, T38V|Y47T had the highest specific activity of all variants with β-leucine followed by S17P|T38V with a specific activity of 1.056 ± 0.008 U µg−1. Also, in case of β-phenylalanine, S17P|T38V had the highest specific activity, followed by T38V|Y47T.
Taken together, the present study shows that several biochemical properties of StoTA could be successfully improved by a rational design based on a previously proposed signature sequence motif for aminotransferases converting aromatic β-amino acids. Not all modifications lead to a higher enzyme activity, but 7 out of 17 variants (S17P, T38V, Y47T, N70E, S17P|T38V, T38V|Y47T and S17P|T38V|Y47T) showed a higher specific activity with at least one substrate. Among all variants, three modifications, Y47T, S17P|T38V and T38V|Y47T not only had, depending on the amino donor, an approx. 4–10.5 times higher activity than the wild type, the same mutants also had a lower optimal temperature of 40–50 °C. These properties make these three variants valuable candidates for industrial processes. Especially, S17P|T38V is a very promising modification, it has the highest specific activities with β-homophenylalanine, β-homoalanine, 4-aminopentanoic acid and β-phenylalanine of all tested variants and can be used to produce these amino acids. Additionally, S17P|T38V has a temperature optimum of only 40 °C, which is 20 °C lower than for the wild type, which makes this variant even more suitable for industrial processes. Only for the production of β-leucine, T38V|Y47T would be better because of the roughly 30% higher specific activity compared to S17P|T38V.
Besides the improvement of the catalytic activity, T317 could be identified as an essential residue. Without this residue, the enzyme loses most of its activity. The assumption that T317 is important for PLP binding is strengthened by the fact that PLP is not added to the model if the T317V variant is folded in silico by swiss-model (https://swissmodel.expasy.org/interactive) (Waterhouse et al. 2018), even if the unmodified structure, which includes PLP, is used as template.
Comments (0)