
Remember me


In this study, four approaches to developing new plant protection products based on targeted chitosan modification were implemented, namely: controlled production of chitosan with defined molecular weights; preparation of chitosan nanoparticle dispersions; synthesis of chelate complexes with biologically active metals; and formation of adducts with organic biologically active substances.
The polysaccharide nature of chitosan and the presence of reactive functional groups in it provide a wide possibility for obtaining diverse chemical modifications, allowing the imparting of new properties to this polymer in accordance with the set task. At the same time, it should be taken into account that special methods of producing chitosan from chitin extracted from various natural sources (krill, fungi, insects, etc.) cause structural and functional heterogeneity of chitosan macromolecules across many parameters and complicate polymer standardization during comparative evaluation. In this regard, this study was performed using the initial chitosan with a molecular weight of 150 kDa and a degree of deacetylation of 85% (Bioprogress, Russia), with which the following types of modification were carried out to enhance biological activity:
Depolymerization of Initial Chitosan under the Action of Chemical Reagents“Natural,” or more precisely, commercially available chitosan, has a molecular weight exceeding 150 kDa, as well as a certain molecular weight distribution. Based on general principles, an understanding of the nature of high-molecular-weight substances, and knowledge of the conformational behavior of polymers, it can be assumed that the biological activity of chitosan—which is determined by the interaction of charged macromolecules with the surface of bacterial or fungal cells—tends to decrease with increasing molecular weight. This is due to the fact that a significant portion of the polymer’s functional groups will be hidden within the volume of macromolecular coils and inaccessible for interaction. On the other hand, a decrease in molecular weight will lead to a weakening of the cooperative effect of the impact of a large number of functional groups belonging to one chitosan molecule due to a reduction in their number. Therefore, one can assume the existence of some optimal molecular weight of chitosan for its use as a biocidal agent, and this molecular weight is likely not to be too high.
Oxidative depolymerization of the initial chitosan was performed in the presence of NaNO2 to obtain samples with molecular weights of 6.5–100 kDa and with the same degree of deacetylation—85%. These samples indeed exhibited different biological activity against the bacteria P. syringae, E. сarotovora, B. polymyxa (Table 1) and the fungi F. oxysporum, S. sclerotiorum (Table 2). Both high- and low-molecular-weight chitosan exhibited noticeably lower bacteriostatic activity. Fungicidal activity depends less on molecular weight. But even this example confirms the existence of a nonlinear dependence of the biocidal activity of chitosan samples on molecular weight.
Table 1. Antibacterial activity of chitosan at different molecular weightsTable 2. Antifungal activity of chitosan samples with different molecular weights in vitroA general pattern in the action of chitosans on all test cultures was revealed. Chitosan with Mw from 5 to 50 kDa possessed sufficiently high antibacterial activity. Reducing the polymer Mw to 3 kDa reduced the inhibitory effect of chitosan on the growth of bacterial test cultures. Chitosan with Mw 150 kDa possessed significantly lower bactericidal activity. E. carotovora exhibited the greatest sensitivity to chitosan polymers. Another Gram-negative test culture, P. syringae, on the contrary, turned out to be the most resistant to the action of the polycation. This indicates that differences in cell wall type between Gram-negative and Gram-positive bacteria do not determine cell sensitivity to the polycation.
All chitosan samples exhibited high fungistatic activity, inhibiting the mycelial growth of F. oxysporum by 44.4–80.0%, and S. sclerotiorum by 60.6–86.3% over 10 days of cultivation compared to the control. The effect of inhibiting fungal colony growth depended on the Mw of chitosan. Low-molecular-weight chitosan (Mw 3 kDa) restrained mycelial growth of the fungus S. sclerotiorum up to 75.5%, but less effectively suppressed the growth of the ascomycete F. oxysporum, inhibiting mycelial growth by only 44% compared to the control. Conversely, high-molecular-weight chitosan (150 kDa) more strongly restrained the growth of the test culture S. sclerotiorum. Chitosans with molecular weights from 5 to 50 kDa equally effectively inhibited the growth of colonies of both fungi over 10 days of cultivation (Table 2).
Thus, the antifungal activity of chitosan at an optimal degree of deacetylation (85%) is determined by its molecular weight, as well as by the type of pathogen.
It was established that all chitosan samples possessed high antibacterial and antifungal activity. As the Mw increased from 3 kDa, the biocidal activity gradually increased, was maximal at Mw 10–50 kDa, and then decreased. Thus, for chitosan, a nonlinear dependence of biocidal activity on its molecular weight is observed. At the same time, it is noteworthy that the efficacy of chitosan against bacteria depended on the phytopathogen species.
Formation of Nanoparticles by Methods of Ionotropic Gelation and Fractional PrecipitationChitosan is a weak cationic polyelectrolyte with a rigid polysaccharide chain. The solubility of chitosan is associated with its amino groups, which, in a protonated state, provide sufficient hydrophilicity. In neutral aqueous solutions, chitosan amino groups are not charged; therefore, the polymer is insoluble. Significant solubility occurs in acidic environments (pH < 6). This can be a limitation for the use of chitosan as a plant protection product due to the negative influence of the acidic environment on the protected object. A solution may be the use of relatively safe acids or those with their own biological activity, such as succinic, benzoic, or salicylic acid.
An alternative approach is to formulate chitosan not as a molecular solution, but as a stable dispersion of nanoparticles. Such dispersions can be prepared in neutral aqueous media without the need for added acid.
Two methods were used to prepare chitosan nanoparticles:
1. Ionotropic gelation, which essentially represents the controlled formation of an insoluble salt (interpolyelectrolyte complex) of chitosan with polybasic acids (inorganic, organic, or polymeric). Under carefully selected conditions (concentration, pH, dosing rate), the formation of an insoluble complex leads to nanoparticle formation. An additional controlling factor can be the introduction of a surfactant into the composition. In our work, for ionotropic gelation, the reaction between a chitosan solution in acetic acid and sodium tripolyphosphate was used. Chemically, the product was chitosan tripolyphosphate (a salt); morphologically, it was a nanoparticle dispersion.
2. Direct precipitation of chitosan from an acidic solution upon its titration with alkali. In this case, a slow increase in the pH of the solution leads to a gradual decrease in the number of ionized amino groups. Upon reaching a certain critical ratio of ionized/non-ionized groups, the chitosan macromolecule loses solubility, collapses, and a new phase is formed. Control of concentration, mixing regime, temperature, and titration rate allows for obtaining neutralized chitosan in the form of a stable nanoparticle dispersion. The advantage of this approach is the possibility of obtaining a stable dispersion (suspension) of nanoparticles of neutral chitosan. Table 3 presents the particle sizes of chitosan dispersions (Chit-5), determined by dynamic light scattering (DLS) performed on a Photocor Complex spectrometer at 25°C at a wavelength λ0 = 445 nm.
Table 3. Hydrodynamic diameters of particles of chitosan samples (Chit-5) obtained by the alkaline precipitation methodChitosan nanoparticles obtained by both methods exhibited significantly higher antibacterial activity compared to simple chitosan taken in the form of a solution at the same concentration (Table 4).
Table 4. Antibacterial and fungistatic activity of chitosan nanoparticles compared to initial chitosanaUsing the agar diffusion method, an assessment of the direct antagonistic action of dispersions Chit-5 and Chit-TPP against test cultures A. solani, as well as C. michiganensis (Table 4), was conducted. The obtained results were compared with the action of a solution of the initial chitosan in acetate buffer (pH 5).
Comparison of inhibition zone diameters showed that the dispersions, regardless of chitosan Mw, possessed greater antibacterial activity than the initial chitosan. The Chit-5 dispersion also possessed fungistatic activity, inhibiting the growth of the fungus A. solani, whereas the solution of the initial chitosan showed zero activity. Note that the fungistatic and bactericidal activity, assessed for Chit-5 dispersions after their storage for three months at +4°C, remained virtually unchanged.
Formation of Chelate Complexes with Variable Valence Metal IonsAnalysis of chitosan chelate complexes with metal ions (Cu, Zn, Fe) revealed general patterns in the manifestation of biological activity depending on the type of metal, molecular weight of chitosan, and type of microorganism. At the same time, the chitosan complex with copper ions turned out to be the most effective, regardless of the magnitude of molecular weight and type of pathogen.
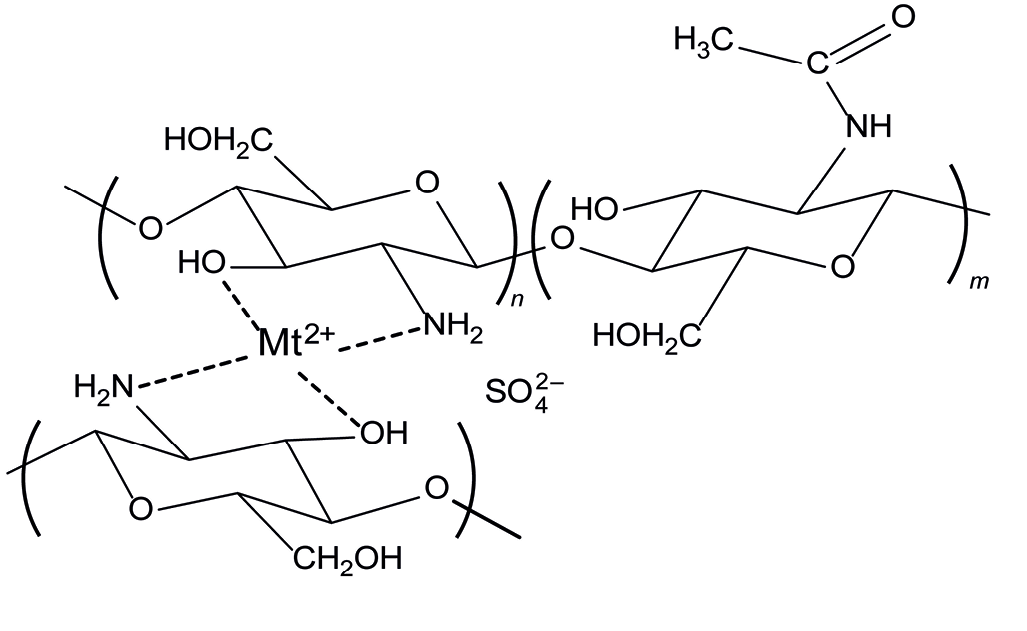
The abundance of amino groups in chitosan enables the formation of chelate complexes with metal ions. Many metals, such as copper, zinc, and iron have pronounced biological activity. Copper(II) complexes are part of many plant protection products. We obtained complexes of chitosan with copper and zinc (Fig. 1).
Fig. 1.
Schematic representation of a chitosan–metal chelate complex with a divalent metal sulfate (Mt2+).
Copper(II) and zinc(II) easily form stable chelate complexes involving chitosan amino groups. Iron does not typically form amine complexes; therefore, to obtain chitosan doped with iron(II), it was necessary to preliminarily introduce functional groups for iron binding into the chitosan molecule structure (Table 5).
Table 5. Metal content in complexes with chitosanFor this purpose, chitosan modification with p-hydroxybenzaldehyde (BA) followed by reduction of aldimine bonds with sodium borohydride was carried out (Fig. 2).
Fig. 2.
Introduction of phenolic groups into chitosan via reaction with hydroxybenzaldehyde for effective binding of iron ions.
Analysis of chitosan chelate complexes with metal ions (Cu, Zn, Fe) revealed general patterns in the manifestation of biological activity depending on the type of metal, molecular weight of chitosan, and type of microorganism (Table 6). Iron (II) ions did not exert a significant influence on the growth of microorganisms, since iron is one of the biogenic elements. The chitosan complex with copper ions was the most effective regardless of the magnitude of chitosan molecular weight and the type of phytopathogen.
Table 6. Antifungal activity of chitosan samples and their complexes with copper, zinc, and ironWhen studying the antifungal activity of chitosan samples against F. oxysporum, it was established that the inclusion of copper and zinc in chitosan contributed to an increase in its antifungal activity. Unlike copper and zinc, iron does not increase the antifungal activity of chitosan.
As a result of chelation with metal ions, the positive charge of chitosan increases, which enhances the adsorption capacity of the polycation and promotes stronger binding of the chitosan molecule to the surface of microbial cellular structures, leading to disruption of their normal metabolism.
Thus, the obtained chitosan complexes with metals (copper and zinc) more effectively suppress the mycelial growth of the studied test fungus.
Table 7 shows that the inclusion of copper and zinc into chitosan increased the antibacterial activity of the initial polymer by 2–3 times.
Table 7. Antibacterial activity of chitosan samples and their complexes with copper and zincaThus, the inclusion of copper and zinc into chitosan with the formation of a chelate complex increased the inhibitory activity of the samples regardless of the Mw magnitude and type of microorganism.
Introduction of Biologically Active Substances into the Chitosan StructureIonic and covalent binding of chitosan with biologically active substances. A key feature of chitosan is its ability to induce biochemical pathways in plants, activating defense reactions and conferring resistance to fungal, bacterial, and viral diseases [14, 15]. That is why preparations based on chitosan should possess the properties of disease resistance inducers and, thereby, contribute to increasing plant productivity.
To enhance the biological activity of chitosan, reactions of its modification with biologically active substances are carried out; these include various organic derivatives, for example, carboxylic acids, phenolic compounds, aldehydes, etc. The introduction of these compounds into the chitosan molecule is possible through ionic or covalent binding. It is known that aromatic acids, such as benzoic, ortho-hydroxybenzoic, salicylic, and p-hydroxybenzoic, exhibit properties of biologically (physiologically) active substances and can improve the properties of chitosan as a resistance inducer.
Salicylic acid (SA) is widely recognized as a key component of the defense signaling system involved in systemic acquired resistance. Markers of this process development are pathogenesis-related (PR) proteins, and SA acts as a signal in the induction of their genes [16, 17].
Benzoic acid (BA)—a precursor of salicylic acid—and para-hydroxybenzoic acid (pHBA), which is a structural analogue of SA, can also be distinguished. There are reports that exogenous BA increases rice resistance to helminthosporiosis (causative agent—Bipolaris oryzae Shoem. (= Helminthosprium oryzae Br. et Haan)), as well as the resistance of cocoa plants to the basidiomycete Ceratobasidium theobromae (= Oncobasidium theobromae, Thanatephorus theobromae) [18, 19]. Such possible action of pHBA has been practically unstudied.
Chitosan samples containing ionically bound aromatic acids were obtained: benzoic acid (Chit+BA); salicylic acid (Chit+SA); para-hydroxybenzoic acid (Chit+PBA) (Fig. 3).
Fig. 3.
Schematic representation of a chitosan complex with salicylic acid.
The immunomodulatory activity of chitosan samples containing ionically bound aromatic acids are presented in Table 8.
Table 8. Influence of chitosan, benzoic acids, and corresponding salts on wheat resistance to tan spot and leaf rustIt was established that chitosan containing SA (Chit+SA) reduced wheat brown rust infection 2.0 times more effectively compared to pure chitosan. High immunomodulatory activity of the chitosan salt with BA, close to the effectiveness of Chit+SA, was revealed, which was expressed in a 60% reduction in wheat leaf infection relative to the control. The inclusion of SA into chitosan also increased its inducing activity against wheat dark brown spot, but less effectively compared to brown rust.
Evaluation of the biological activity of the salts Chitosan-BA and Chitosan+pHBA showed that as resistance inducers against dark brown spot, they were less effective than the initial chitosan (25% leaf infection) and the Chitosan+SA variant. In these experimental variants, the area of leaf infection by the ascomycete C. sativus was 60 and 50%, respectively.
One of the least understood aspects of phytoimmunity is the mechanism regulating immune response formation during infection by pathogens with different nutritional strategies. However, it is precisely the solution to this task, associated with the ability to direct metabolism by changing gene expression in the desired direction, that ultimately determines the possibility of controlling the pathological process [20, 21].
Numerous studies of the molecular mechanisms of phytoimmunity have led to the understanding that the formation of a defensive response to pathogen invasion is ensured by changes in cell metabolism and activation of the plant’s genetic apparatus, and the intensity of the organism’s immune responses can be regulated by exogenous biologically active substances—immunomodulators [15, 22, 23].
It is assumed that, depending on the pathogen’s nutritional strategy, plants, in response to infection, activate different signaling systems to ensure optimal protection of their tissues.
Based on their nutritional strategy, pathogens are classified as biotrophs, necrotrophs, and hemibiotrophs. When plants are infected by pathogenic fungi, the most significant defensive reaction is the increased generation of reactive oxygen species (ROS) [24]. Under these conditions, processes of necrosis formation are triggered, leading to the death of the biotroph if the host plant is resistant to the pathogen. At the same time, unlike the obligate pathosystem, a favorable environment compatible for the necrotroph is formed [25]. It follows that increased ROS generation is effective against biotrophic pathogens, since their functioning requires living host plant cells. In the case of a necrotrophic type of parasitism, the strategy for increasing resistance should be aimed at inactivating ROS and inhibiting necrosis formation processes and stimulating the cellular antioxidant system [26]. Therefore, these pathogen nutritional strategies should be considered when formulating an effective plant protection strategy. To date, there are insufficient works in this direction, although the problem itself is very relevant. Thus, when wheat is infected by a biotroph and a hemibiotroph, defensive reactions are triggered, the basis of which is a certain balance between the level of ROS and the antioxidant potential in the plant. Whether oxidative stress will progress or be blocked will directly affect the nature of the relationships developing between the pathogen and the host plant [27].
Thus, during biotrophic infection, it is necessary to increase ROS levels to protect plants, whereas in the case of a hemibiotroph, on the contrary, it is desirable to reduce the level of ROS in the cell to slow down the development of necrosis and limit the spread of the pathogen.
In accordance with this approach, the developed disease resistance inducers should be compounds with multidirectional functions (hybrid) and contain two active centers in their structure: one will promote ROS generation, the other, conversely, will inactivate it. At the same time, it is important to observe the temporal period of release of the introduced low-molecular components from the system, which can be determined by the type of chitosan-BAS bond.
For creating such hybrid systems, chitosan is the most promising. A large number of reactive groups allows for covalent or ionic immobilization of biologically active substance molecules on the chitosan macromolecule, thus supplementing its own biological activity with the specific action of the corresponding modifier. Due to the large number of amino groups per one chitosan molecule, it is possible not only to vary, within certain limits, the concentration of the modifier, but also to combine several biologically active substances in one composition. Thus, the chitosan macromolecule can act as a platform for combining substances with different mechanisms of action.
As BAS with pro-oxidant properties, the classic inducer of disease resistance—salicylic acid—was used, which plays a central role in plant defense against biotrophic pathogens [28, 29].
To obtain chitosan samples possessing antioxidant activity (AO) (hereinafter referred to as conjugates), vanillin was chosen—a known natural and synthetic compound potentially possessing AO properties due to the presence of a sterically hindered phenolic group in its structure. Vanillin was attached to chitosan with the formation of Schiff bases (aldimine bonds), which were then reduced with an excess of sodium borohydride.
The attachment of an acid to the chitosan macromolecule by an ionic bond predetermines their high mobility, and hence the biological effect in the short term. On the other hand, covalent binding of biologically active substance molecules—vanillin—ensures a slower action.
Hybrid polymeric systems, Chit-Van+SA, were developed, representing chitosan containing simultaneously covalently attached vanillin (25 wt %) and ionically bound salicylic acid, the amount of which was varied relative to the attached vanillin from 1 : 0.5 to 1 : 2 (Fig. 4).
Fig. 4.
Schematic representation of a molecule of a combined preparation containing ionically bound salicylic acid and covalently bound vanillin.
The results of a comparative evaluation of the influence of chitosan and its derivatives with Van and SA on wheat resistance to pathogens of different trophic types showed that chitosan containing ionically bound SA (Chit+SA) is more effective in protecting plants from the biotroph P. recondita, whereas the conjugate (Chit-Van) more effectively induces resistance to the hemibiotroph B. sorokiniana. Experimentally, it was revealed that for the manifestation of high inducing activity in relation to the biotroph P. recondita and the hemibiotroph C. sativus, the molar ratio of Van and SA in hybrid immunomodulators should be within 1 : 1–1 : 2. Such a ratio of low-molecular-weight BAS was optimal and allowed for the targeted regulation of the plant immune response to infection, sequentially activating plant defensive reactions.
According to literature data, the inducing effect of exogenous SA in increasing plant resistance to phytopathogens is due to its ability to inhibit catalase—an enzyme that detoxifies hydrogen peroxide, leading to the accumulation of ROS [30, 31]. The latter are considered important signaling mediators in the formation of plant resistance. Considering the literature and our obtained data [32], it can be assumed that the high inducing activity of hybrid systems (Chit-Van+SA), in which the Van : SA ratio is 1 : 1 or 1 : 2, is associated with the ability of exogenous SA to be the first to be released from the polymeric system and induce H2O2 generation to an amount that leads to the activation of a whole spectrum of plant defensive reactions inhibiting pathogen development. This increases the duration of the asymptomatic biotrophic stage of hemibiotroph development and significantly slows down disease progression. Increased peroxide content during this period also contributes to the formation of resistance to the biotroph, and the development of P. recondita is reduced to 10–15% of the control (Table 9).
Table 9. Influence of chitosan and its derivatives with Van and SA (concentration 0.1% by chitosan) on wheat resistance to pathogens of different trophic typesThese findings suggest that the effectiveness of disease resistance inducers is determined by their ability at the stage of pathogen invasion to change the conditions of its development in the plant by regulating redox processes, which are fundamental in the dynamics of pathogenesis.
A new approach to creating resistance inducers based on chitosan containing fragments with antioxidant and pro-oxidant activities in a certain ratio has been proposed. Chitosan modified simultaneously with salicylic acid and vanillin (Fig. 4) represents an optimal variant of a polymeric system with regulated immunomodulatory properties for protecting wheat plants from pathogens with different types of nutrition.
Thus, as a result of the work performed, it has been established that chitosan, as a natural biologically active polymer, represents not so much a ready-to-use product, but rather a multifunctional and promising material for developing on its basis a wide range of polyfunctional antibacterial and antifungal agents with combined activity and diverse mechanisms of action.
Comments (0)