
Remember me



A 61-year-old Caucasian female with a medical history of Vater papilla adenocarcinoma in complete remission was admitted for FLAG-M salvage chemotherapy (fludarabine 50 mg/day on days + 1 to + 5, cytarabine 1700 mg/day on days + 1 to + 5, mitoxantrone 10 mg/day on day + 1, and 15 mg/day on days + 2 to + 3) due to relapsed acute myelomonocytic leukemia (AMMoL), which had previously transformed from adverse-risk chronic myelomonocytic leukemia (Table 1). On day + 7 following chemotherapy initiation, the patient developed chemotherapy-induced aplasia. By day + 22, she presented with new-onset febrile neutropenia, requiring the initiation of empirical ceftazidime.
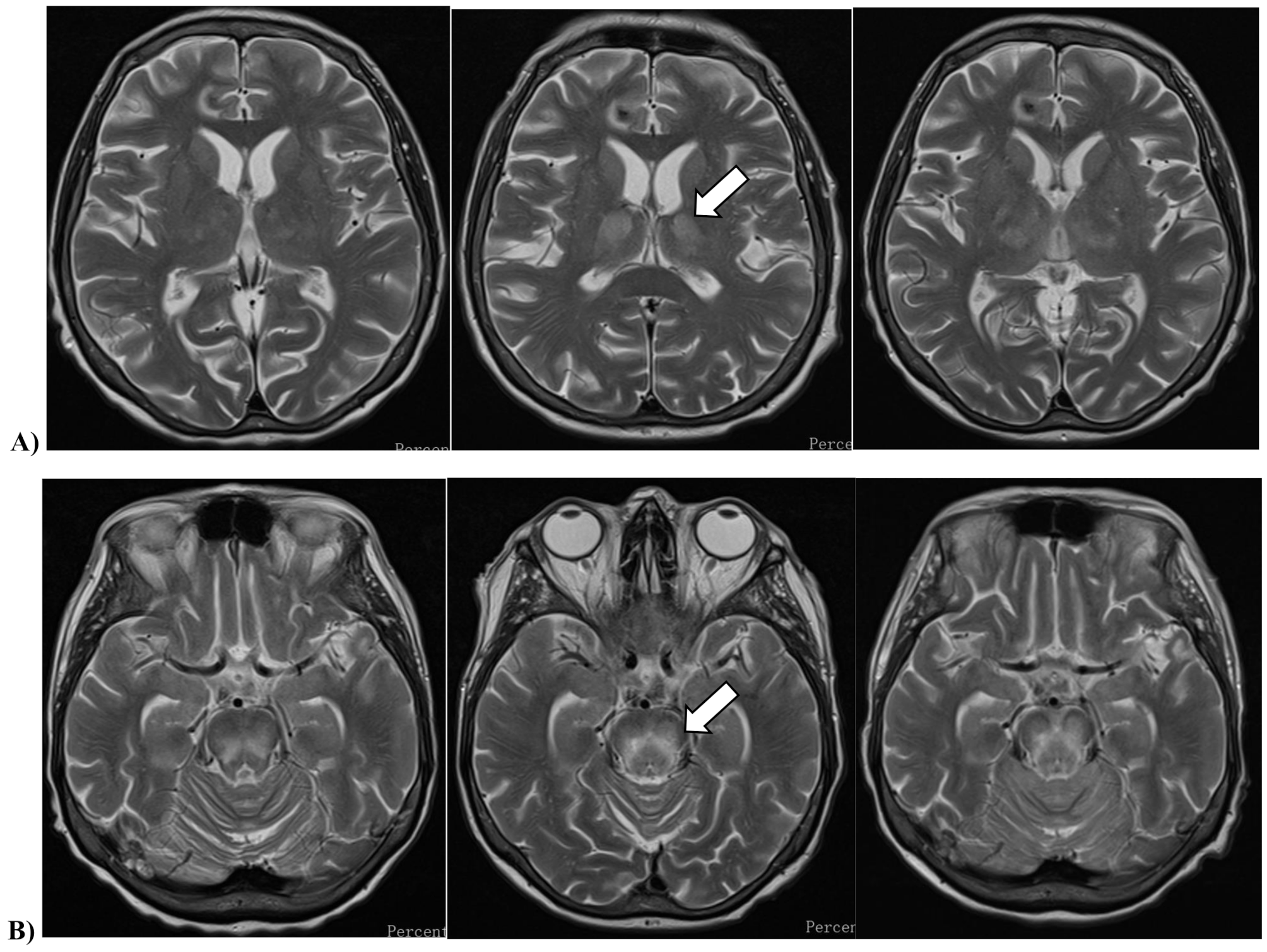
Table 1 Clinical characteristicsOn day + 25, the patient developed new-onset diplopia, prompting an urgent cranial magnetic resonance imaging (MRI), which revealed symmetric abnormalities in the thalamus and brainstem, suggestive of metabolic encephalopathy. Multiple small foci in the brain parenchyma were consistent with microhemorrhages, though the morphology also raised suspicion of infectious foci. Differential diagnoses included Wernicke encephalopathy and osmotic demyelination syndrome (Fig. 1). A cerebrospinal fluid (CSF) analysis was unremarkable, but an elevated Aspergillus sp. DNA load of 471 copies/mL by polymerase-chain reaction (PCR) and a concurrently positive serum galactomannan antigen index of 3.0 by chemiluminescence immunoassay prompted for empirical antifungal therapy with liposomal amphotericin-B. Routinely used microbiological diagnostic kits for different pathogen targets are listed in the Supplementary File 1. All tests yielded negative results, with the exception of Aspergillus sp. A chest and paranasal CT scan was also done with unremarkable result.
Fig. 1
Cranial magnetic resonance imaging of the patient, showing focal progression with time. T2-weighted turbo spin echo sequences, acquired in the transverse planes of A) the thalamus and B) the midbrain
On day + 29, the patient developed new-onset dysarthria and somnolence, with clear neurological deterioration. A repeat cranial MRI demonstrated significant progression of extent and intensity in the previously observed symmetric signal abnormalities in the thalamus, cerebral peduncles, brainstem tegmentum, and pons. Additionally, smaller signal abnormalities with mild progression were observed in the medulla oblongata and the craniospinal junction. A lumbar puncture was repeated, with CSF samples sent for routine bacterial/fungal culture and Gram smear, multiplex PCR for bacterial, viral and fungal pathogens, as well as CSF/blood samples Aspergillus sp., Cryptococcus sp. and Toxoplasma gondii (Supplementary File 1.). In addition, a dissemination of the underlying AMMoL was ruled out by CSF flow cytometry. Additional CSF, blood, and urine samples were sent to the National Center for Public Health and Pharmacy for testing of WNV, USUV, tick-borne encephalitis (TBEV), and JC virus (JCV) (Table 2.).
Table 2 Results of the molecular biological assaysThe patient became nearly unresponsive with tachypnea and episodes of desaturation the next morning, prompting an intensive care unit (ICU) admission. A chest X-ray revealed diffuse patchy opacity in the lungs, suggestive of an infectious infiltrate. During the day, persistent desaturation and secretion retention necessitated endotracheal intubation and mechanical ventilation, while intravenous antibiotics were escalated to meropenem and levofloxacin. An extensive microbiological testing for healthcare-acquired pneumonia from bronchoalveolar lavage was done by routine bacterial/fungal culturing and Gram smear, multiplex PCR for bacterial and fungal pathogens as well as Pneumocystis jirovecii, Aspergillus sp. and CMV, with a repeated set of blood cultures (Supplementary File 1).
Given the suspicion of WNV infection due to extreme seasonal activity in the region, off-label empirical antiviral therapy with remdesivir and intravenous immunoglobulin (IVIG) was initiated. On the second day of ICU admittance, the National Center for Public Health and Pharmacy reported that the patient’s CSF, urine, and blood samples tested positive for Usutu virus (Table 2.). A follow-up cranial MRI revealed slight progression of previously described symmetric signal abnormalities in the thalamus and pons, while EEG showed moderate background detected in the theta range, with desynchronization in response to external stimuli, without any focal abnormalities or seizure activity. A repeat neurological consultation recommended continued supportive care, and intracranial pressure-reducing therapy was deemed unnecessary. Microbiological tests led to the discontinuation of remdesivir, and processes were initiated for the acquisition of favipiravir. Despite all efforts, the patient died on day + 37. Autopsy revealed histopathological signs of diffuse microglial activation with incipient gliosis in the brain, and diffuse alveolar damage in the lungs. Apart from USUV, no other infectious agents affecting the central nervous system (CNS) could be documented by extensive microbiological or pathological diagnostics.
Virological investigationLaboratory diagnosis of human arbovirus infections in Hungary is centralized and carried out exclusively at the National Reference Laboratory for Viral Zoonoses at the National Center for Public Health and Pharmacy, according to the decree of the Ministry of Health 18/1998 (VI. 3.) [21]. Serum, anticoagulant-treated whole blood, CSF, and urine samples were investigated by molecular diagnostic methods. Manual nucleic acid extraction was carried out by the QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer’s recommendations. Tick-borne encephalitis virus (TBEV), WNV, and USUV RNA were detected by reverse-transcription real-time PCR (RT-qPCR) method. For the WNV RNA detection, RealStar® WNV CE-IVD RT-PCR Kit 2.0 (Altona Diagnostics, Hamburg-Altona, Germany) was used. TBEV and USUV RT-qPCR were performed according to in-house developed protocols based on primer and TaqMan probe sequences published elsewhere [22, 23]. The adapted in-house RT-qPCR protocols were validated prior to use with clinical specimens. The intra-assay precision was 0.78% coefficient of variation (CV) for the TBEV RT-qPCR assay and 0.69% CV for the USUV RT-qPCR assay. The limit of detection (LoD) for the TBEV RT-qPCR assay was established at a Ct value below 40.00. The USUV RT-qPCR assay demonstrated a LoD at a Ct value of 35.00, corresponding to a limit of quantification of 4 copies/µL of USUV RNA. Amplification efficiency ranged from 106 to 111%, according to the original study by Nikolay et al. [23]. Further differential diagnosis included testing for polyomavirus DNA using the GeneProof BK/JC Virus PCR Kit (GeneProof, Brno, Czech Republic). The results of the molecular biological investigation are summarized in Table 2. Despite the patient’s immunocompromised status, serological tests were also performed to detect TBEV, WNV, and USUV antibody response in the serum and CSF samples. For the detection of TBEV and WNV-specific IgG, IgM, and IgA antibodies, the Flavivirus Mosaic 1 (EUROIMMUN Medizinische Labordiagnostika, Lübeck, Germany) indirect immunofluorescence assay (IFA) was used. USUV-specific IgG/IgM/IgA antibody response was examined using an in-house developed IFA method. In-house USUV immunofluorescence tests were performed as described earlier [24]. Both serum and CSF samples were negative for TBEV, WNV, and USUV-specific IgG, IgM, and IgA antibodies.
Virus isolationIn vivo virus isolation from the USUV-positive post-mortem brain specimen was performed by intracranial inoculation of newborn mice. On day 7 post-infection, mice were euthanized, and after dissection and preprocessing, the mouse brain was homogenized using MagNA Lyser Instrument (Roche Diagnostics, Basel, Switzerland). The USUV RT-qPCR result confirmed virus replication in the mouse brain, as a lower Ct value (Ct 14.29) was measured compared to the clinical specimen (Ct 26.94; Table 2.). In addition, the supernatant of the mouse brain homogenate was inoculated onto Vero E6 (ATCC; CRL-1586) cell culture. Cells were cultured for up to 7 days, and the cytopathic effect was monitored daily. USUV RT-qPCR was carried out from the cell culture supernatant (passage 2 isolate) and confirmed the USUV replication (RT-qPCR Ct value of 14.87).
Sequencing and phylogenetic analysisUSUV RT-qPCR results were confirmed by amplicon-based whole-genome next-generation sequencing (NGS) method. NGS was performed on samples from two patients with confirmed USUV infection in 2024, along with USUV-positive mosquito pools collected during the same period. Clinical specimens from the two patients were sent for NGS to the Bernhard Nocht Institute for Tropical Medicine, Hamburg, Germany. The NGS of mosquito pools was conducted at the National Reference Laboratory for Viral Zoonoses (Hungary). To run an amplicon-based sequencing of subsequent mosquito pools, a multiplex primer panel was designed using the Primal Scheme online tool (https://primalscheme.com). Primer sequences are listed in the Supplementary File 2. NGS was also performed on the USUV passage 2 isolate, using sequence-independent amplification of the viral RNA in this case [25].
The DNA library preparation was performed using the Nextera XT DNA Library Preparation Kit (Illumina, San Diego, CA; USA), following the manufacturer’s instructions. Sequencing was carried out on the Illumina MiSeq System (Illumina, San Diego, CA; USA) with MiSeq Reagent Micro kit, v2, 300 cycles (MS-103-1002) (Illumina, San Diego, CA, USA). For the NGS data preprocessing, assembly, and reference mapping, the Geneious Prime (Version 2025.1) software was used. The nearly whole-genome sequences were submitted to the NCBI GenBank database. The name of the USUV strain (passage 2 isolate) is USUV-HUN-SM2024, and the GenBank Accession number is PQ772639. Based on GenBank records, detailed information on all USUV sequences obtained during the 2024 transmission season in Hungary is summarized in Supplementary File 3.
Phylogenetic analysis was based on nearly whole-genome sequences of the USUV strains. Pairwise alignment of the viral sequences was performed using Nucleotide BLAST (Basic Local Alignment Search Tool; https://blast.ncbi.nlm.nih.gov/Blast.cgi). For phylogenetic analysis, multiple sequence alignment was conducted using MAFFT (Multiple Alignment using Fast Fourier Transform; version 7: https://mafft.cbrc.jp/alignment/server/index.html). The phylogenetic tree was constructed with MEGA11 (Molecular Evolutionary Genetics Analysis, version 11). Maximum Likelihood analysis was performed under the General Time Reversible model with gamma-distributed rate variation among sites and a proportion of invariant sites (GTR + G + I). Bootstrap support values were estimated using 1,000 replicates.
Usutu virus phylogeny and spread in Hungary, 2024The co-circulation of different USUV lineages in Europe has been well documented in recent years. To date, at least eight distinct phylogenetic lineages have been described: three African and five European lineages (EU1–EU5) [26]. However, the terminology for lineages is not yet fully standardized; for instance some studies refer to EU5, while others define it as the Middle Eastern clade [27]. Among the European lineages, the EU2 is the most prevalent in Central and Eastern Europe, including Austria, Germany, Hungary, Italy, and Romania [28, 29]. A study by Zecchin et al. revealed that in Northeastern Italy, USUV strains clustered into two distinct groups, designated as EU2-A and EU2-B sublineages. According to phylogeographic analysis, Italy played a key role in the spatial dissemination of European USUV lineages. For instance, the Emilia-Romagna region has been identified as a major source of viral spread to Austria and Germany [29]. Two introduction events from Austria to Hungary have been identified: one in the 1990s and another between 2013 and 2016 [27, 29]. The EU2-A sublineage is considered to have high neurovirulence and has been dominant in recent outbreaks [30].
The virological differential diagnosis of WNV and USUV infections led to the identification of three laboratory-confirmed human USUV cases during the 2024 transmission period in Hungary. All three cases were associated with severe neurological manifestations. Symptom onset for the first case (Patient No. 1) occurred in early August, while the other two were diagnosed in September (Patient No. 2) and early October (Patient No. 3), respectively. The geographical distribution of the human USUV infections was relatively dispersed (Fig. 2). The presumed place of exposure for Patient No. 3 (presented in this study) is Budapest, as the patient was hospitalized at the National Institute of Hematology and Infectious Diseases during the incubation period. Between May 9 and November 8, 2024, as part of a pilot study, mosquito collection and pathogen surveillance were carried out at seven trapping sites in and around the capital city of Budapest (Fig. 2). A total of 7,410 female Culex spp. mosquitoes were collected and tested for USUV in 474 pools. USUV was detected in eight mosquito pools from five of the seven trapping sites. The first positive results were obtained from sampling sites No. 4 and No. 5 on July 21, while the last USUV-positive pool was identified at site No. 6 on August 25, 2024. All sampling sites and the locations of human exposure are illustrated in Fig. 2. USUV RNA was detected in clinical samples from Patients No. 1 and No. 3. Whole genome sequencing and subsequent phylogenetic analysis were conducted on all eight mosquito-positive pools, as well as the USUV-positive human specimens. Although mosquito surveillance was limited to a well-defined area, the combined evidence from mosquito and human infections demonstrates an active USUV circulation in Hungary. The resulting phylogenetic tree, shown in Fig. 3, revealed that Hungarian sequences clustered within European lineage 2.
Fig. 2
Geographical locations of confirmed human USUV infections and mosquito sampling sites in 2024. Whole genome USUV sequences are available from Patient No.1 and Patient No.3 (marked in red). Mosquito collection and USUV screening were carried out in Budapest. The locations of mosquito trapping sites, where USUV RT-qPCR pools were identified are marked with purple dots. The place of exposure of the immunocompromised patient (Patient No.3) is marked with a light blue dot. The map was created using the online map editor geojson.io (Retrieved June 15, 2025, from https://geojson.io/#map=12.91/46.90257/17.6427).
Fig. 3
Maximum likelihood phylogenetic tree of nearly complete genome sequences of USUV strains. Sequence data are labeled with the following information: GenBank accession number, geographical location of virus isolation or detection, year of isolation or detection, and the scientific name of the host species. Sequences corresponding to the presented case are marked with light blue dots, while a dark blue diamond indicates sequences from another human case and eight mosquito pools collected in 2024
In the 2024 Hungarian USUV sequences, two amino acid substitutions were identified: G595S in the E protein and E3425D in the NS5 protein; both mutations are characteristic of the EU2-A sublineage [28, 29]. Additional mutations specific to Italian strains [29] were also found in all Hungarian sequences: E830G (NS1 protein), I1324T (NS2A protein), and M2645I (NS5 protein). These findings are consistent with the phylogenetic analysis and confirm the close genetic relationship between Hungarian and Italian USUV strains (Fig. 3.). Notably, the G595S substitution is located within the DIII domain of the envelope protein and may play a role in the neuroinvasive potential of USUV in humans [31]. Most human neurological cases have been associated with the EU2 lineage, particularly the EU2-A sublineage, suggesting a potential link between the genetic characteristics and neurotropism [15]. However, this assumption has some limitations, such as the relatively small number of human cases and human-derived sequences, as well as the lack of detailed clinical data. Overall, the currently circulating EU2-A USUV strains in Hungary may pose a risk to human health, especially during periods of intense mosquito activity, which could lead to an increased number of human infections.
Comments (0)