
Remember me

The unique properties of intrinsically disordered proteins (IDPs), namely a high degree of structural and dynamic heterogeneity, as well as the presence of largely solvent-exposed backbones, have a significant impact on NMR observables, influencing the quality of NMR spectra and the information that can be retrieved. Due to the absence of a well-defined 3D structure and the dynamic interconversion among various conformers, most contributions to chemical shifts are averaged out, resulting in crowded spectra. This is reflected in a severely limited proton chemical shift range, with significant degeneracy of signals. Consequently, standard NMR experiments may not provide sufficient resolving power for unambiguous resonance assignment, especially in the case of primary sequences with highly repetitive amino acid motifs. On the other hand, manipulating selective pulses for IDPs is more straightforward because signals cluster around random coil values for each amino acid, with minor deviations influenced by neighboring amino acids, and amide and aliphatic protons hence fall in two distinct spectral regions, well separated from each other.
Increasing resolution: N-TROSY with selective decoupling on aliphatic 1HThe use of selective adiabatic pulses on aliphatic protons during N-TROSY experiments can help to avoid the evolution of long-range couplings between 1Hali and 15N nuclear spins. A specific Constant Adiabatic Hyperbolic Secant pulse (CaPowHsec) was employed in a decoupling scheme designed to cover the aliphatic 1H region (approximately 3.5 ppm) during 15N signal acquisition. The pulse had a bandwidth of about 3.5 ppm, with sweep width set to 3.4 kHz at 800 MHz proton Larmor frequency and a pulse length of 8 ms.
The features of the CaPowHsec pulse were first tested on 1D 1H NMR spectra of α-synuclein. A simple modification of the excitation sculpting pulse sequence, replacing both the hard and water-selective π pulses with CaPowHsec π pulses selective for aliphatic protons, yields spectra showing only the aliphatic protons, illustrating the selectivity of the chosen CaPowHsec pulse (Figure SI1).
Fig. 1
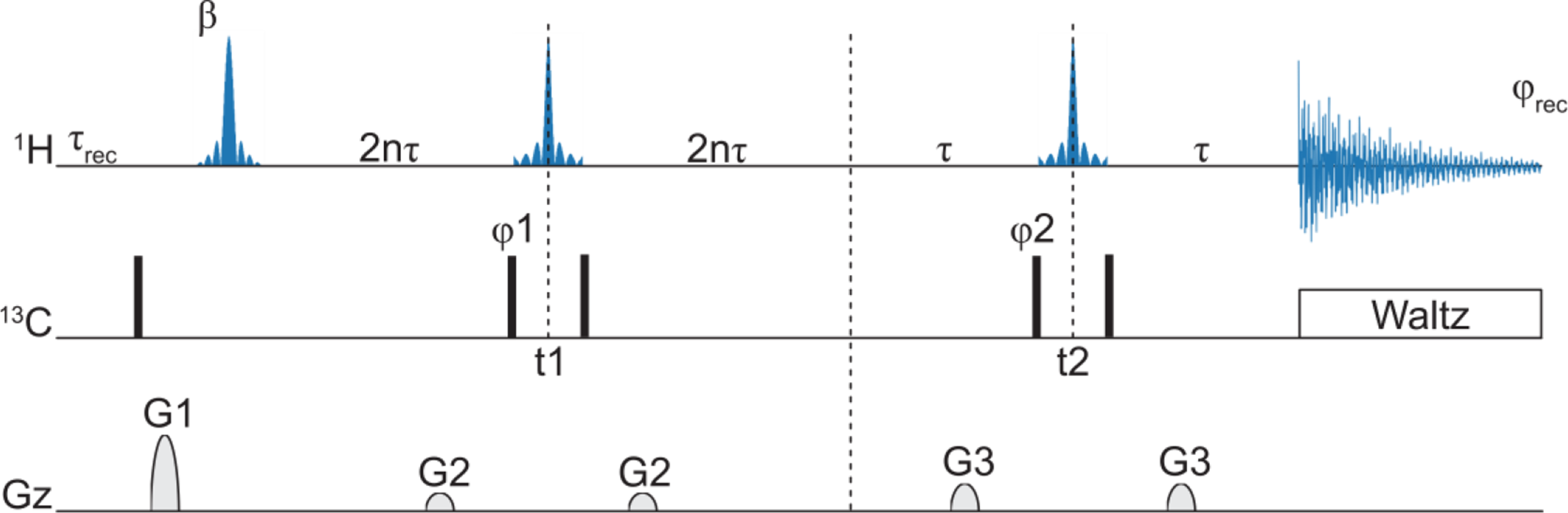
Panel a illustrates the overlay of the 800 MHz 15N detected TROSY spectra acquired on a sample of α-synuclein (1.0 mM, 20 mM potassium phosphate at pH 5.5, 200 mM NaCl, 0.05 mM EDTA, 0.05% NaN3, 0.02% Protease Inhibitor Cocktail) at 283 K. The spectrum with selective decoupling of 1Hali (red) is compared to the one acquired without decoupling (blue). The comparison shows a decrease in 15N linewidth of the signals, which can be appreciated in the zoomed spectral regions shown in panel b. The line narrowing effect is smaller in glycine signals (panel c). A schematic representation of the N-TROSY pulse sequence is reported in panel d, accordingly with the already published pulse sequence (Takeuchi et al. 2016). Narrow and wide black rectangular shapes represent π and π/2 pulses, the phases are φ1 (y -y x -x) and φrec (x -x -y y). The delay τ was set to 2.7 ms (1/4JHN). The gradient g1 and g2 were applied with a strength of 31% and 19% respectively. A CaPowHsec3.4,8.0,1.0.8 adiabatic pulse was used to selectively decouple the 1H aliphatic region
We then used this CaPowHsec pulse in the context of 1H composite pulse decoupling (CPD) during 15N acquisition in an N-TROSY experiment on α-synuclein. To assess the effective increase in resolution obtained by aliphatic 1H decoupling, a control N-TROSY experiment was recorded with the same acquisition parameters, but without 1Hali decoupling. The data acquired at 283 K are reported in Fig. 1 and show that decoupling leads to a decrease in 15N linewidth of 42% on average along the entire backbone (see Fig. 2a). Similar behavior is observed when the experiments are repeated at higher temperatures, with average decreases in linewidth due to 1Hali decoupling of 47% and 44% at 293 K and 303 K, respectively (Figure SI2).
Careful analysis of the spectra shows that the average decrease in linewidth is less pronounced (around 16%) when focusing on signals from glycine residues as shown in Fig. 1c. It is worth noting that glycine residues, lacking a 1Hβ proton, are characterized by an intrinsically narrower linewidth (4.1 Hz on average at 283 K) compared to non-glycine residues (5.7 Hz on average at 283 K). Both linewidth and signal-to-noise analysis are reported only for well-isolated peaks, to avoid introducing errors due to partial signal overlap.
The benefits of using 1Hali decoupling are also confirmed by the resulting increase in signal-to-noise ratio (S/N) of the peaks, with an average increase of 26% at 283 K. Indeed, when a decoupling scheme is applied, peaks split due to the J coupling should collapse to singlets, contributing to an increase in signal intensity. This effect is clearly visible in Fig. 2b where nearly all signals gain in sensitivity. An exception is represented again by glycine residue signals, which show a mild decrease in S/N when the decoupling is applied. With increasing temperature, this tendency is extended to other residues (serine, threonine, and histidine) whose 1HN are characterized by a high exchange rate with water protons (Figure SI2).
Fig. 2
The effect of 1Hali decoupling on linewidth and signal-to-noise ratio is illustrated in this figure. Panel a depicts the 15N linewidths measured on a sample of α-synuclein for the 15N-detected TROSY acquired at 283 K. Solid bars represent the experiment without decoupling, while open bars show the linewidth values for the experiment with decoupling. Solid and dashed horizontal lines indicate average linewidth values for experiments with and without decoupling, respectively. The signal-to-noise ratio values measured for the experiments with decoupling (open bars) and without (solid bars) are reported in panel b. Only well-isolated peaks are reported in this graph to minimize potential sources of error. Additionally, the data from proline residues are not present, because of their lack of amide protons
In the N-TROSY experiments presented so far, the 1H decoupling frequency carrier was set to 3.0 ppm in order to fully cover the aliphatic region. 1D 1H experiments as shown in Figure SI1 had however indicated that the CaPowHsec pulse perturbs water proton magnetization if it is centered at this frequency. To evaluate the impact of this perturbation, an N-TROSY spectrum was acquired with the 1H decoupling frequency carrier set to 2.0 ppm, where water magnetization is unaffected (Figure SI1). The N-TROSY experiments with 1H decoupling frequency carrier at 2.0 and 3.0 ppm showed similar performance, suggesting negligible influence of water proton magnetization perturbation (see Figure SI3). We thus kept the decoupling frequency carrier at 3.0 ppm in subsequent experiments for optimal coverage of the aliphatic proton. To test the experiments on CBP-ID4, we used a 1.2 GHz spectrometer. When the magnetic field increases, so does the required bandwidth in Hz. Consequently, the spectral region to be covered is larger, but the spectral separation (in Hz) between amide and aliphatic protons also increases. The advantage of increased separation of resonances due to higher field is less crucial in the case of IDPs, since the resonances are already in well-separated regions. On the other hand, the wider spectral window to be covered does not seem to be a limitation for the selected proton decoupling. To record the N-TROSY experiments, the CaPowHsec pulse was designed to cover the region of aliphatic protons (about 3.5 ppm), differing from the previous one only for the sweep width, which was set to 5.1 kHz. The experiments conducted at 298 K, reported in Fig. 3, indicate consistent performance compared to the experiments acquired at lower field on α-synuclein. Indeed, the linewidth analysis shows an average decrease of 41% when decoupling is applied, as reported in Figure SI4. The performance remains less impressive in glycine residues, with an average decrease in linewidth of 18%, a tendency confirmed also by the S/N trend.
Fig. 3
Panel a illustrates an overlay of 15N detected TROSY acquired with and without decoupling on a 1.2 GHz spectrometer, on a sample of CBP-ID4 (around 340 µM, 25 mM sodium acetate at pH 5.5, 50 mM NaCl, 2 mM TCEP, 0.05 mM EDTA) at 298 K. The spectrum with selective 1Hali decoupling is shown in red, the one without it in blue. The comparison exhibits a decrease in signal linewidth, which can also lead to resolving previously overlapped peaks. In panel b for example, the signals arising from Q30, Q54 and Q179 are well separated when 1Hali decoupling is applied. As already observed in α-synuclein, the linewidth decrease is less pronounced for signals of glycine residues (panel c). Panel d illustrates the effect of 1Hali decoupling on the 15N transverse coherence. Fourier transform of the indirect dimension for the N-TROSY without decoupling is shown in blue, with a maximum time during which signals are still visible lower than 200 ms. In red, the same data is presented for the experiment with decoupling, with an observed increase in the detection time of about 300 ms. Both experiments were acquired and processed with comparable parameters
Of course, improvements in linewidth are only visible when the Free Induction Decay (FID) is acquired long enough to benefit from the long-living coherence, at least for 300 ms in the present case, as qualitatively illustrated in Fig. 3d. The presence of decoupling plays in fact a key role in enhancing the detection time of the 15N magnetization. When decoupling is not active, the FID can be observed for less than 200 ms, while the CaPowHsec decoupling enables a longer detection of the magnetization, around 300 ms.
The long coherence lifetimes typical of IDPs permit to increase the chemical shift evolution in the indirect dimension. The J couplings between Hali and HN are operative during this period contributing to an increase in linewidth resulting from (unresolved) homonuclear scalar couplings. To further increase the resolution of N-TROSY experiments, it is possible to refocus the evolution of these couplings inserting one band selective pulse on Hali during the chemical shift evolution of these couplings in the indirect dimension as shown in Figure SI5.
Better sensitivity with the BEST approachWhen observing heteronuclear spins such as 15N, which is characterized by a low gyromagnetic ratio (γ), sensitivity might be an issue. For experiments that exploit 1H as starting polarization source, band-selective irradiation of a subset of 1H nuclear spins accelerates magnetization recovery (Solyom et al. 2013, Lescop et al. 2007, Schanda et al. 2009, Lescop et al. 2010), enabling shorter recovery delays which in turn allow to either reduce the duration of an NMR experiment or to accumulate more transients in the same amount of time, with a net result of increased S/N (Lescop et al. 2010). An important consideration for proteins in which amide protons experience efficient chemical exchange with water protons consists in maintaining the latter unperturbed during the experiment, to avoid compromising longitudinal relaxation enhancement. This is the base of the BEST approach irradiating only a subset of nuclear spins, such as the imino-protons in RNA (Schnieders et al. 2017).
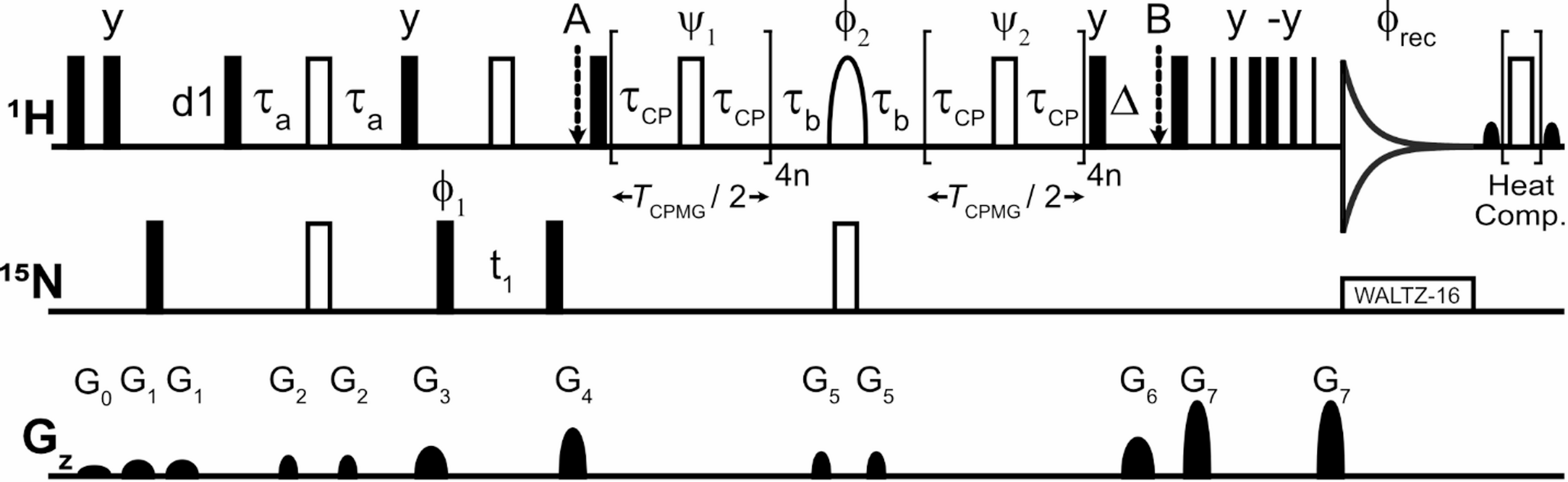
Here we present a variant of the BESTN-TROSY experiment for application to IDPs. In this version, the proton pulses (π and π/2 pulses) were designed to selectively excite and refocus the amide region, ensuring minimal interference with the water magnetization. The chosen pulse shapes were Reburp.1000 and Eburp2.1000 or Eburp2tr.1000 (Geen and Freeman 1991) for π and π/2 pulses, respectively, designed to cover 3.0 ppm and centered at 9.0 ppm. This pulse configuration was carefully selected paying attention to the frequency ranges of their sidebands, to avoid perturbation of the water resonance as much as possible.
To assess the performance of 1H longitudinal relaxation enhancement by the BEST approach, the experiment with selective pulses was compared with the one with hard pulses on the proton channel. These experiments were performed on both α-synuclein and CBP-ID4 samples. In all cases, a constant acquisition time of 350 ms was used, to fully exploit the longer lifetime of 15N nuclear spin magnetization, while the recycle delay between the end of the acquisition and the beginning of the following experiment was progressively reduced. Thus, the sum of detection time and recycle delay, named recovery time (Figure SI6), for BEST experiment was 1350 ms. Then, it was shortened to 675 ms, 450 ms, and 355 ms, reducing the experimental duration almost by a factor of 4. Figure 4a illustrates the average peak volume per unit time measured for BESTN-TROSY experiments recorded on α-synuclein as the recycle delay is varied, at three different temperatures (blue 283 K, green 293 K, orange 303 K). All experiments were acquired with the same number of scans using a spectrometer operating at 800 MHz.
To assess the performance of 1H longitudinal relaxation enhancement by the BEST approach, the experiment with selective pulses was compared with the one with hard pulses on the proton channel. Figure 4a also reports the measured average volume for the experiment with hard pulses on 1H acquired by using a recovery delay of 1350 ms. The advantage of using selective pulses on amide protons for application to IDPs/IDRs is striking. Comparing two experiments with the same recovery delay (1350 ms) at 283 K on α-synuclein, there is a 29% increase in measured peak volume when the experiment is acquired with the BEST principle. In the BEST-type experiment, the sum of detection time and recycle delay can be reduced while increasing signal per unit time. The plot also highlights the different behavior when temperature is changed. The initial part of the coherence transfer pathway of both experiments involves 1HN magnetization, and this is reflected in a reduction in sensitivity when the sample is in conditions of more effective exchange of amide protons with water. A similar trend is observed for CBP-ID4. The one-dimensional projections of the 15N dimension (Fig. 4c) confirm the efficacy of selective irradiation, with an average intensity gain of 27% for identical recycle delays. To identify the optimal recovery delay for maximum sensitivity per unit time, the average S/N ratios per unit time can be plotted as a function of time, as shown in Figure SI6.
Fig. 4
Average measured cross peak volumes per unit time at three different temperatures measured on α-synuclein for BESTN-TROSY experiments varying the recovery time (1350 ms, 675 ms, 450 ms and 355ms) are presented in panel a. Blue solid squares represent data at 283 K, green points at 293 K, and orange triangles at 303 K. Empty symbols refer to average volume per unit time for N-TROSY experiment. Panel b illustrates the one-dimensional projection of the 2D 15N-TROSY spectra with BEST (blue) and without (green) acquired on α-synuclein at 800 MHz and 283 K using identical acquisition times, recycle delays, and number of scans. The same comparison is reported in panel c for CBP-ID4 acquired at 298 K using a spectrometer operating at 1200 MHz. In both experiments, an increase in sensitivity is observed. The pulse sequence is reported in panel d according to the already published pulse sequence (Schnieders et al. 2017). The black narrow and wide rectangular shapes represent hard π and π/2 pulses while the rounded ones represent π and π/2 Reburp.1000 and Eburp2.1000 shaped pulses. the phase cycle is represented by φ1 (y -y x -x) and φrec (x -x -y y). The delay τ was set to 2.7 ms (1/4JHN). The gradient g1 and g2 were applied with a strength of 31% and 19% respectively. A CaPowHsec adiabatic pulse with a sweep-width of 3.4 kHz was used to selectively decouple the 1H aliphatic region at 800 MHz while at 1200 MHz the sweep width was increased to 5.1 kHz
Combining strategiesIn the previous sections, the potential resolution enhancement achievable through selective decoupling in the aliphatic proton region was demonstrated, as well as the increase in sensitivity resulting from exploiting the longitudinal relaxation enhancement. Here, we combine the two strategies in a BESTN-TROSY experiment, using both selective amide proton excitation and aliphatic proton decoupling via CaPowHsec pulses during acquisition. The frequency carrier of the latter pulses was set to 2.0 ppm, to selectively cover the aliphatic region without perturbing the water magnetization. Figure 5a illustrates the measurable reduction in linewidth achieved on CBP-ID4 spectra with the application of decoupling and BEST approach using a recycle delay of 300 ms respect to the experiment acquired with the same recovery delay but without the decoupling. The comparison demonstrates an average decrease in linewidth of approximately 44%, similar to N-TROSY spectra. On the other hand, the increase in sensitivity as measured by signal-to-noise values (Fig. 5b and c) was not as pronounced as expected, resulting in comparable values in the two experiments. It is worth noting that the region of the protein most negatively affected by the presence of decoupling, in terms of S/N, is between residues 101–128 where there is a higher propensity for α-helix formation (Piai et al. 2016). The presence of decoupling perturbs the aliphatic protons and thus reduces the LRE effects arising from dipolar cross-relaxation rates which are larger for regions with higher structural propensity (see Figure SI7).
Fig. 5
The effects on linewidth and signal to noise of 1Hali decoupling in BESTN-TROSY spectra for CBP-ID4 are reported in the figure above. Panel a illustrates the decrease in linewidth when 1Hali decoupling is applied. Light green bars depict the linewidth (Hz) of signals acquired in a 15N detected BESTN-TROSY without decoupling. Solid dark green bars represent the linewidth of signals obtained from the same experiment with selective decoupling applied to aliphatic protons during acquisition. The dashed orange line indicates the average linewidth measured for the experiment without decoupling, while the dotted-dashed orange line illustrates the same value for the experiment with decoupling. Panel b displays the signal to noise ratio measured for the experiment without decoupling, whereas panel c shows signal to noise values for the experiment with decoupling. The differences between the values in panels b and c are plotted in Figure SI7. For this graph, only isolated peaks were analyzed to minimize potential errors. Furthermore, data from proline residues are absent since they lack an amide proton
Thus, while additional gains in sensitivity from using aliphatic proton decoupling in the context of BESTN-TROSY experiments appear to depend on secondary structure propensity, the combined use of both BEST-type selective amide proton excitation and aliphatic proton decoupling allows for increases in resolution. This strategy can be fruitfully exploited to observe particularly crowded regions of proteins, and it can complement the already promising resolution performance due to the use of high or ultra-high magnetic fields and 15N direct detection.
It is also possible to identify the optimal recovery delay for experiments that exploit combined strategies to implement faster measurements with an optimal signal-to-noise (S/N) ratio per unit time (Figure SI6). The experiments were conducted on α-synuclein at 1.2 GHz. The detection time was kept constant at 350 ms, while the recycle delay varied from 5 ms to 2 s.
Comments (0)