
Remember me



Quantifying the infectivity of pathogens is a crucial step toward the understanding of infection risks. Human challenge trials provide unique opportunities to understand infectious dose, disease progression, and immune response for various pathogens.1 Their advantage, in addition to field trials, has been recognized before and during the COVID-19 pandemic.2,3 Human challenge studies are often used for evaluating the clinical efficacy of novel interventions (e.g., antiviral drugs and vaccines), measured as a reduction in infection risk between control and treatment arms. In most trials, the infection risk is observed as the proportion of infected participants among those exposed to a single dose. However, as infection risk is dose dependent, this setup may not fully capture the efficacy of interventions.
We can study how the infection risk depends on the challenge dose, using data from multiple challenge studies with different doses, or ideally from multiple-dose studies. Observed dose–response relationships are described by various formulations (i.e., dose–response models).4,5 Dose–response models can be used to improve trial designs,6 to describe infectivity and immunogenicity in human hosts,7 to assess the infection risks at lower exposure settings (e.g., indoor environment),5 and to simulate the infection risks via various transmission routes.8 They are also used to find optimal inoculation dose levels9 to test interventions, but the interventions themselves are almost always tested with a single inoculation dose.
Dose–response models can account for individual variation in susceptibility among participants exposed to the same dose, which is crucial when interpreting the difference in infection risks. Most often, the individual variation has been modeled by a continuous distribution, such as a beta distribution10,11 or gamma distribution.12,13 However, when a proportion of individuals is completely immune, the variation in susceptibility is better captured by other distributions (e.g., a bimodal distribution14). When conducting challenge trials for endemic diseases, participants often had been exposed to the pathogen of interest before the trial.
Here, we start by reformulating dose–response models with a flexible description of the variation in host susceptibility that allows for an intuitive biological interpretation. We show how variation in susceptibility determines the dose–response relationship for endemic diseases where the study population may be partially immune, using challenge data on the endemic human coronavirus HCoV-229E. As an example of applying the proposed method to novel pathogens (i.e., pathogens to which the study population is fully susceptible), we compute the plausible range of SARS-CoV-2 dose–response curves based on available outcomes of a challenge study.
Our approach suggests how the design of human challenge trials can be improved to better capture the variation in susceptibility and suggests how to translate the outcomes of human challenge studies into infection risks for the general population.
METHODS Human Challenge StudiesWe conducted a literature search to collect available data from human challenge studies with endemic coronaviruses and SARS-CoV-2. The collected data consists of five studies with endemic coronaviruses HCoV-229E and one study with SARS-CoV-2. In all cases, the study population consisted of healthy adult volunteers, and the participants were intranasally inoculated with certain doses in each trial. The challenge studies reported the challenge dose, the number of challenged individuals, and the number of infected individuals as summarized in eAppendix, https://links.lww.com/EDE/C81 and eTables 1 and 2, https://links.lww.com/EDE/C81.
Dose–response Models to Analyze Human Challenge StudiesThe reported infectious doses d are expectations of a Poisson distribution of the actual infectious dose, with mean d. If each host is equally susceptible, the probability of infection P(d) given a challenge dose d is P(d)=1−exp(−d). This assumes that each infectious particle can independently establish an infection.5,15
We capture the variation in susceptibility among study participants by assigning each a level of susceptibility s, according to a distribution f(s) with mean 1. A participant with a level of susceptibility s has s times higher probability of infection compared with an average individual, per infectious particle. The probability of infection upon challenge with a dose d is P(d)=1−∫0∞exp(−sd)f(s)ds. By fitting this model for the probability of infection to the observed proportion of infections in human challenge studies, we can infer the shape of the distribution f(s) using the method of maximum likelihood (see eAppendix, https://links.lww.com/EDE/C81, for details). We use four different models for the distribution: a Dirac delta distribution to reflect a situation where all individuals have the same level of susceptibility; a gamma distribution to reflect a situation where the level of susceptibility varies continuously; a bimodal distribution with one fraction of the population almost immune, and the remaining fraction with a single level of susceptibility; and a bimodal distribution with one fraction of the population almost immune and the remaining fraction with a gamma distribution for the level of susceptibility to vary continuously. Detailed model descriptions and estimated parameters are provided in eTable 3, https://links.lww.com/EDE/C81. All codes and analyzed data are available at the author’s GitHub link (https://github.com/fmiura/CoronaDR_2022).
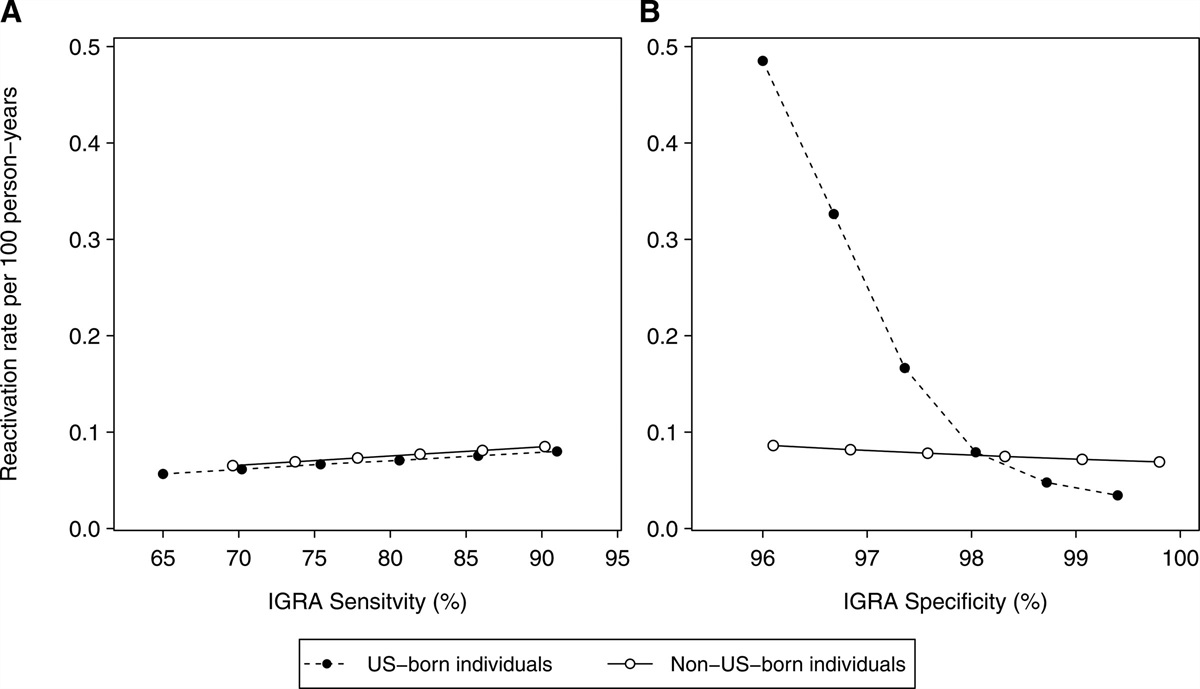
RESULTS Susceptibility Distributions Determine the Shape of Dose–response CurvesWe described the proportion of infections among individuals exposed to different doses of the endemic coronavirus HcoV-229E by fitting dose–response models. Since the collected trial data include participants who might have been exposed to viruses before, we included bimodal distributions of susceptibility. We visualized the flexibility of such distributions and the goodness of fit of those models (Figure 1C,D), while models with homogeneous or continuous susceptibility distributions did not capture the data well (Figure 1A,B).
 FIGURE 1.:
FIGURE 1.: Estimated dose–response models of the endemic coronavirus HCoV-229E based on observed human challenge data. Gray bubbles indicate the observed data, and the size of bubbles indicates the number of participants in each trial. To describe different heterogeneity in susceptibility, dose–response models with delta (A), gamma (B), two-level (C), and gamma with point-mass distributions (D) were fitted to the data.
The results provided strong statistical support for a distribution reflecting a situation where a fraction of the population is almost immune whereas the remaining fraction of the population has a single level of susceptibility. There is no statistical support for a homogeneous level of susceptibility or continuous variation in susceptibility for all individuals (Table 1).
TABLE 1. - Comparison of Estimated Dose–response Models Based on Model Fit to the Observed Endemic Coronavirus Challenge Data Delta Gamma Two-level Gamma+point-mass Log-Likelihood –393.0 –107.2 –98.3 –98.3 No. of parameters 1 2 3 4 AICa 787.9 218.5 202.6 204.6 Difference in AICb 585.3 15.9 0 2.0aThe lowest value indicates the best model in terms of prediction.
bDifference of >10 indicates strong evidence.16 The values here suggest substantial support for the heterogeneity in susceptibility.AIC = Akaike Information Criterion.
In the available human challenge study with SARS-CoV-2, all participants were healthy young adults with no evidence of prior SARS-CoV-2 infection or vaccination, and they were all exposed to the same single dose.17 Here, we show how the variation in susceptibility would affect the infection risk at different doses. Since participants had not had any prior exposure to this virus, we assumed a continuous variation in susceptibility only (i.e., we assumed there is no pre-existing immune fraction in the study population). We fitted the model with several gamma distributions for the level of susceptibility s to the observed SARS-CoV-2 challenge data, where we increased the coefficient of variation (CV) over orders of magnitude from small (10−6) to large (102). The corresponding curves reveal that the infection risk increases more gradually with increasing CV in susceptibility (Figure 2A).
 FIGURE 2.: Dose–response curves of SARS-CoV-2 (A–C) and comparison with an estimated dose–response curve of endemic coronaviruses (D) based on observed human challenge data. In panel A–C, light red dots stand for observations from human challenge data of SARS-CoV-2 (eTable 2, https://links.lww.com/EDE/C81), and whiskers show Jeffrey’s binomial confidence intervals (95%). (A) illustrates that the steepness of fitted curves (blue lines) decreases over the increase in the CV in susceptibility from 10-6 to 102. (B,C) show curves fitted to 1000 bootstrapped samples from observed data where the CV is set as 0 and 1, respectively. These values are selected to consider two scenarios, where the susceptibility level is completely homogeneous and where the level is comparable to that of endemic coronavirus infection. Dotted gray lines show reference dose–response curves of SARS-CoV-1 animal model.18 In (D), plots and whiskers are observed challenge data of endemic coronaviruses (eTable S1, https://links.lww.com/EDE/C81), and dark and light blue lines indicate fitted SARS-CoV-2 models where the CV is fixed as 0 and 1 respectively.
FIGURE 2.: Dose–response curves of SARS-CoV-2 (A–C) and comparison with an estimated dose–response curve of endemic coronaviruses (D) based on observed human challenge data. In panel A–C, light red dots stand for observations from human challenge data of SARS-CoV-2 (eTable 2, https://links.lww.com/EDE/C81), and whiskers show Jeffrey’s binomial confidence intervals (95%). (A) illustrates that the steepness of fitted curves (blue lines) decreases over the increase in the CV in susceptibility from 10-6 to 102. (B,C) show curves fitted to 1000 bootstrapped samples from observed data where the CV is set as 0 and 1, respectively. These values are selected to consider two scenarios, where the susceptibility level is completely homogeneous and where the level is comparable to that of endemic coronavirus infection. Dotted gray lines show reference dose–response curves of SARS-CoV-1 animal model.18 In (D), plots and whiskers are observed challenge data of endemic coronaviruses (eTable S1, https://links.lww.com/EDE/C81), and dark and light blue lines indicate fitted SARS-CoV-2 models where the CV is fixed as 0 and 1 respectively.We accounted for the statistical uncertainty in the proportion of the participants that were infected by taking 1000 bootstrap samples from a binomial distribution with a fixed number of Bernoulli trials (n = 34) and the success probability (P = 18/34) from the original study and fitting dose–response curves to each bootstrapped dataset, reflecting a situation where the susceptibility level is completely homogeneous (CV=0), and where the level is similar to that of the endemic coronavirus infection (CV=1) (Figure 2B,C).
We compared the bootstrapped SARS-CoV-2 dose–response curves with the dose–response curve from a SARS-CoV-1 mouse model obtained by Watanabe et al,18 which has been widely used in risk assessments of SARS-CoV-2 (dotted lines in Figure 2 and eFigure 1, https://links.lww.com/EDE/C81). This reveals that using the current reference model based on mouse data with SARS-CoV-1 could lead to a serious underestimation of the infection risk for SARS-CoV-2, irrespective of the shape of the distribution of susceptibility level. We also compared the SARS-CoV-2 dose–response curves with the observed outcomes from the challenge studies with endemic coronavirus (Figure 2D). This suggests that the estimated range of infection risks of endemic coronavirus is consistent with the observed infection risk in the SARS-CoV-2 trial population.17
DISCUSSIONIn this study, we revealed the plausible range of infection risk over multiple orders of magnitude of the infectious dose for the endemic coronavirus HCoV-229E and SARS-CoV-2, based on human challenge trials. We presented how these dose–response relationships are shaped by the underlying distribution of susceptibility to infection.
The range of SARS-CoV-2 dose–response curves arises from the unknown distribution of background susceptibility in the population and the statistical uncertainty due to the limited number of participants in the human challenge study. Our results caution against assuming equal susceptibility in the population in risk assessments,8,18 as this assumption results in a lower bound for infection risks at lower doses.
Our results provide implications for further research. We address three of them here.
First, our approach suggests a possible improvement in the design of human challenge trials. Conventional trials tend to use a single dose such as the median human infectious dose (HID50).19 Using two (or more) challenge doses would be highly informative for extrapolating the findings over a wider range of doses. If multiple challenge doses would not be feasible, it is advisable to consider using a different challenge dose from those used in previous studies. In fact, many current trials choose the same inoculation dose referring to previous trials.20 However, determining the dose–response curve would be beneficial to those who develop challenge trials, because one can design trials with more precise expectations on the number of infected individuals (or, consequently, the statistical power to test interventions) when escalating or decreasing the dose level. When higher doses are not acceptable from a safety perspective, lower doses are also useful for determining dose–response relationships. The outcomes of different studies can be combined in a meta-analysis that takes advantage of those different doses to infer how infection risk changes with dose. This would elucidate the unknown variation in level of susceptibility among individuals.
Second, the dose–response models proposed here, as many other dose–response models, have underlying assumptions that the infectious particles are homogeneously mixed in the inoculates and act independently in causing an infection.5,15 These assumptions suffice for describing the outcome of human challenge studies, even though it might not hold, for example, when virus particles aggregate. Our dose–response modeling approach can be extended to allow for variation in the per-particle probability, using methods explored previously,5,21 or to incorporate study-specific effects with a Bayesian framework,22 which would allow for a built-in check of violating this assumption. Unlike existing nonmechanistic models, the strength of our formulation lies in its biologic interpretability—we can choose appropriate susceptibility distributions based on evidence from other studies, in addition to the trial data themselves. For example, whereas noroviruses are not able to infect certain individuals due to the lack of binding receptors in the host,6 there is no evidence for such a mechanism for SARS-CoV-2, where binding receptors have only been shown to affect viral load23; hence, our choice for a unimodal distribution for the SARS-CoV-2 challenge data. As challenge trial data are often limited, we recommend incorporating findings from other epidemiologic or clinical experiments when determining the dose–response relationship in a host.
Third, our approach offers guidance for translating the observed risk of infection in human challenge studies to the general population. Currently, this translation is difficult, as the study population of challenge studies consists of healthy adult volunteers for safety and ethical reasons.3 Such a study population is not representative of a general population, which includes children and the elderly. Besides, the general population now includes persons who have been exposed to SARS-CoV-2. The strict selection of healthy adult study participants with no evidence of previous exposure inevitably reduces the variation in susceptibility compared with the general population. The proposed dose–response models with a flexible distribution of susceptibility allow for exploring the impact of an expected increase in variability of the level of susceptibility in the general population. As the variation in susceptibility is expected to be larger in the general population, the dose–response relationship would be flatter. The degree of variation in the general population could be examined with additional observations, such as biomarkers of immunologic protection against infection, if these are measured in both the study participants and the general population. Ultimately, the quantified variation in susceptibility could further inform other epidemiologic transmission models (e.g., Gomes et al24), to explore the impact of interventions at the population level by incorporating the heterogeneity in susceptibility.
In conclusion, our study reveals plausible dose–response relationships for SARS-CoV-2, based on information from human challenge trials, which are consistent with dose–response curves obtained for human endemic coronaviruses. Human challenge trials would be more informative if different doses were used. When extrapolating the observed infection risks in the specific study population to the general population, the expected higher variability of susceptibility in the general population should be taken into account.
ACKNOWLEDGMENTSWe thank Susan van den Hof for reviewing the manuscript.
REFERENCES 1. Adams-Phipps J, Toomey D, Więcek W, et al. A systematic review of human challenge trials, designs, and safety. Clin Infect Dis. 2023;76:609–619. 2. Eyal N, Lipsitch M, Smith PG. Human challenge studies to accelerate coronavirus vaccine licensure. J Infect Dis. 2020;221:1752–1756. 3. Jamrozik E, Littler K, Bull S, et al.; WHO Working Group for Guidance on Human Challenge Studies in COVID-19. Key criteria for the ethical acceptability of COVID-19 human challenge studies: report of a WHO working group. Vaccine. 2021;39:633–640. 4. Furumoto WA, Mickey R. A mathematical model for the infectivity-dilution curve of tobacco mosaic virus: theoretical considerations. Virology. 1967;32:216–223. 5. Haas CN, Rose JB, Gerba CP. Quantitative Microbial Risk Assessment. John Wiley & Sons; 2014. 6. Atmar RL, Opekun AR, Gilger MA, et al. Determination of the 50% human infectious dose for Norwalk virus. J Infect Dis. 2014;209:1016–1022. 7. Clements ML, Snyder MH, Sears SD, Maassab HF, Murphy BR. Evaluation of the infectivity, immunogenicity, and efficacy of live cold-adapted influenza B/Ann Arbor/1/86 reassortant virus vaccine in adult volunteers. J Infect Dis. 1990;161:869–877. 8. Zhang X, Wang J. Dose–response relation deduced for coronaviruses from coronavirus disease 2019, severe acute respiratory syndrome, and middle east respiratory syndrome: meta-analysis results and its application for infection risk assessment of aerosol transmission. Clin Infect Dis. 2021;73:e241–e245. 9. Osowicki J, Azzopardi KI, Fabri L, et al. A controlled human infection model of Streptococcus pyogenes pharyngitis (CHIVAS-M75): an observational, dose-finding study. Lancet Microbe. 2021;2:e291–e299. 10. Teunis PF, Havelaar AH. The Beta Poisson dose–response model is not a single-hit model. Risk Anal. 2000;20:513–520. 11. Gomes MGM, Lipsitch M, Wargo AR, et al. A missing dimension in measures of vaccination impacts. PLoS Pathog. 2014;10:e1003849. 12. King JG, Souto-Maior C, Sartori LM, Maciel-de-Freitas R, Gomes MGM. Variation in wolbachia effects on aedes mosquitoes as a determinant of invasiveness and vectorial capacity. Nat Commun. 2018;9:1483. 13. Langwig KE, Wargo AR, Jones DR, et al. Vaccine effects on heterogeneity in susceptibility and implications for population health management. MBio. 2017;8:e00796–e00717. 14. Halloran ME, Longini IM Jr, Struchiner CJ. Estimability and interpretation of vaccine efficacy using frailty mixing models. Am J Epidemiol. 1996;144:83–97. 15. Nilsen V, Wyller J. QMRA for drinking water: 1 revisiting the mathematical structure of single-hit dose–response models. Risk Anal. 2016;36:145–162. 16. Burnham KP, Anderson DR. Multimodel inference: understanding AIC and BIC in model selection. Sociol Methods Res. 2004;33:261–304. 17. Killingley B, Mann AJ, Kalinova M, et al. Safety, tolerability and viral kinetics during SARS-CoV-2 human challenge in young adults. Nat Med. 2022;28:1031–1041. 18. Watanabe T, Bartrand TA, Weir MH, Omura T, Haas CN. Development of a dose-response model for SARS coronavirus. Risk Anal. 2010;30:1129–1138. 19. Levine MM, Abdullah S, Arabi YM, et al. Viewpoint of a WHO advisory group tasked to consider establishing a closely-monitored challenge model of coronavirus disease 2019 (COVID-19) in healthy volunteers. Clin Infect Dis. 2021;72:2035–2041. 20. Roestenberg M, Hoogerwerf M-A, Ferreira DM, Mordmüller B, Yazdanbakhsh M. Experimental infection of human volunteers. Lancet Infect Dis. 2018;18:e312–e322. 21. Teunis PFM, Brienen N, Kretzschmar MEE. High infectivity and pathogenicity of influenza A virus via aerosol and droplet transmission. Epidemics. 2010;2:215–222. 22. Thebault A, Teunis PFM, Le Pendu J, Le Guyader FS, Denis J-B. Infectivity of GI and GII noroviruses established from oyster related outbreaks. Epidemics. 2013;5:98–110. 23. Loo L, Waller MA, Moreno CL, et al. Fibroblast-expressed LRRC15 is a receptor for SARS-CoV-2 spike and controls antiviral and antifibrotic transcriptional programs. PLoS Biol. 2023;21:e3001967. 24. Gomes MGM, Ferreira MU, Corder RM, et al. Individual variation in susceptibility or exposure to SARS-CoV-2 lowers the herd immunity threshold. J Theor Biol. 2022;540:111063.
Comments (0)