
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Remember me
Limbs' movements result from the complex interaction between brain centers, the spinal cord, and the musculoskeletal system [45]. The spinal network is essential in the control, coordination, and modulation of locomotion [32]. While there is direct evidence of a central pattern generator (CPG) in mammals and other vertebrates [1, 32], the lack of direct experimental access in humans means that there is only indirect evidence [39]. Furthermore, sensory feedback pathways may play a major role compared to other mammals, and lower vertebrates in humans [24–26, 28, 34]. Different studies suggested that the muscle activity observed during human locomotion may be controlled by five locomotor primitives that could be generated by rhythmic neural circuits [14, 30]. Investigating these questions about the roles of different spinal components in controlling locomotion is challenging since we possess only partial knowledge of the interactions between the different subsystems involved in this process. On top of that, limited experimental access complicates the observation of the sub-components functions leading to difficulties in model validation. Computer simulations are necessary and have been proven useful in the past to evaluate the contribution of each control component by evaluating different models, and parameters [3, 18, 19, 22, 29, 41, 48].
Various neuromechanical models have been proposed in the past to address these questions. In 1995, Taga proposed a musculoskeletal system controlled by a neural rhythm generator (RG) composed of seven pairs of neural oscillators and simple sensory-motor signals [49]. Successively, in 2001, Ogihara and Yamazaki developed a neural controller composed of motoneurons (MNs) receiving inputs from a common CPG and reflexes from stretch and force receptors, where the spindle reflexes had inhibitory inputs to antagonist's muscles [40]. In the context of locomotion controlled by CPG mechanisms, Aoi et al (2010) constructed a CPG model based on a two-layered hierarchical network composed of a RG and a pattern formation (PF) layer. The RG model produced rhythmic information using phase oscillators and was regulated by phase resetting based on foot-contact gait events, whereas the PF model generated feedforward commands composed of five motor primitives based on the muscle synergies analysis performed by Ivanenko et al [3, 30]. On the other hand, Geyer and Herr demonstrated that the kinematics and muscle activation observed in human locomotion could be reproduced without CPG commands by relying purely on sensory feedback activated at specific gait cycle phases [22]. The activation of sensory feedback was regulated by a state-machine mechanism (i.e. a set of if–then–else rules) that enabled and disabled specific reflexes depending on the detection of stance or swing phases. A similar controller with partial modifications has then been proposed by Ong et al [41]. In these studies, the activation of sensory responses in the gait cycle is regulated by a state-machine mechanism activating and deactivating reflexes in five gait cycle subphases. The necessity of including such state-machine mechanisms in reflex-based controllers hints at the need for a more sophisticated circuit that controls the underlying reflexes. Other studies have integrated CPG commands on top of these purely sensory-based controllers, showing the benefits of rhythmic circuits in gait modulation [19, 53].
In this study, we propose a novel bio-inspired controller composed of a feedforward network inspired by Aoi et al consisting of two CPGs that produce the locomotor rhythm and patterns and a new physiologically plausible implementation of spinal reflexes based on neurophysiological studies in locomotion [54, 56] without relying on any state machine mechanism. This network controls nine muscle actuators generating torques in a previously assessed musculoskeletal model [17].
The performance of this controller in replicating the behavior of human locomotion and its modulation are investigated and compared with previous experimental and neuromechanical studies. In addition, we investigate the performance of the sensory feedback controller alone to verify whether it is possible to generate human walking behavior with a purely reflex-based controller without relying on state machine mechanisms and to verify the benefit of CPG mechanisms. Finally, we examine the contribution given by pattern generation and reflex circuits to the MNs at slow, intermediate, and fast speeds performing a correlation analysis to identify possible parameters responsible for speed modulation. With these experiments, we aim to address the following questions:
What is the role of CPGs and reflex circuits in the generation of muscle activation in human locomotion?Can low-level feedback circuits produce stable locomotion without a CPG or state-machine?Is the contribution of these two neural components changing with increasing of gait speed?Our results show that the reflex rules implemented in previous models [22, 41] could be reproduced into less abstract and more realistic models of neural circuits. The insights given by the proposed controller suggest that spinal reflexes alone could not reproduce rhythmic locomotion without a state machine mechanism regulating the activation of reflexes in specific phases of the gait cycle. CPG networks appear to play the role of state machines in previous models and to be necessary to promote muscle activation in specific gait cycle phases. In addition, the modulation of CPG frequency seems necessary to modulate step duration. The modulation of either reflexes, CPG network, or both could generate gaits in a wide speed range, highlighting the high level of versatility of the neurospinal control of human locomotion.
This study used the Simulated Controller OptimizatioN Environment (SCONE) software simulation framework [21]. SCONE is an open-source software performing predictive simulations of biological motion, allowing the investigation of individual models and control parameters on the motion. To perform a predictive simulation in SCONE, it is necessary to run a scenario including the following components:
A mechanical model of the system to simulate.A controller that generates input for the model actuators.An objective that describes the target task to optimize, through a weighted combination of measures (sub-components of the cost function).An optimizer that optimizes the free parameters in a scenario for a specific objective.In this study, SCONE software was extended to implement and optimize the new spinal model generating gait simulations of 10 s.
2.1. Musculoskeletal modelThe musculoskeletal model (figure 1) has the skeletal structure presented by Delp et al [17] with a height of 1.8 m and weight of 75.16 kg. The model is constrained in the sagittal plane and has a total of nine degrees of freedom (DoFs): a three-DoFs planar joint between the pelvis and the ground and three rotational DoFs per leg: hip flexion/extension, knee flexion/extension, and ankle dorsiflexion/plantarflexion. Three spheres per leg are used as contact models with the ground: one of radius 5 cm at the calcaneus and two of radius 2.5 cm at the toes. The musculoskeletal model is actuated by nine Hill-type muscle-tendon units per leg: gluteus maximus (GMAX), biarticular hamstrings (HAMS), biceps femoris short head (BFSH), rectus femoris (RF), iliopsoas (ILPSO), vasti muscle group (VAS), gastrocnemius (GAS), soleus (SOL), and tibialis anterior (TA).
Figure 1. Musculoskeletal model used to study human locomotion. The model is constrained in the sagittal plane and has nine DoFs: hip and knee flexion/extension, ankle plantar/dorsal flexion for each leg, and a three-DoFs planar joint between the pelvis and the ground. Movements are generated by the activation of nine muscles per leg: gluteus maximus (GMAX), biarticular hamstrings (HAMS), biceps femoris short head (BFSH), rectus femoris (RF), iliopsoas (ILPSO), vasti (VAS), gastrocnemius medialis (GAS), soleus (SOL), and tibialis anterior (TA).
Download figure:
Standard image High-resolution image 2.2. ControllerMuscle activation is regulated by the excitation provided by the MNs. The MNs are stimulated or inhibited by the different components of the bio-inspired controller: the balance controller of the trunk and the spinal network, composed of the CPGs and spinal reflexes. The balance controller and the CPGs are modeled at an abstract level. Indeed, the former provides balance inputs in specific phases of the gait cycle with proportional derivative (PD) controllers, and the latter is composed of two abstract oscillators generating primitive patterns. By contrast, the spinal reflexes are modeled more realistically at a lower level of abstraction and are structured in different leaky integrator neurons divided into three types: somatosensory neurons (SNs), interneurons (INs), and MNs. The overall structure of the controller is reported in figure 2. The balance controller of the trunk regulates only the activation of hip muscles in specific phases of the gait cycle, whereas CPGs and spinal reflex controllers provide inputs to all muscles and are not regulated by any state-machine mechanism. We chose to maintain the state machine for the balance controller in order to simplify the balance control since our main goal is the simulation of locomotor movement. A physiological neuromechanical model of trunk balance control is a complex task that is outside the scope of this study. Muscle excitation is triggered by the MN output with values between 0 and 1 since MNs can only provide excitation to muscle fibers and cannot have negative outputs. To keep a reasonable level of abstraction and complexity, we will assume that the neuron's output (corresponding to its firing rate)  follows the dynamics of a leaky integrator:
follows the dynamics of a leaky integrator:
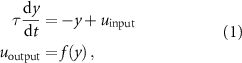
where y is the neuronal response,  is the neural input, τ the time constant (typically 0.01),
is the neural input, τ the time constant (typically 0.01),  the output of the neuron, and f the activation function. The activation function used for MNs is the min–max operator (
the output of the neuron, and f the activation function. The activation function used for MNs is the min–max operator ( ), and the neural input is defined as:
), and the neural input is defined as:
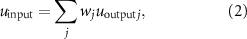
where wj is the weight associated with the jth connection, and  the output of the jth neuron. The MN receives inputs from the CPGs' network (
the output of the jth neuron. The MN receives inputs from the CPGs' network ( ) and the reflex circuit (
) and the reflex circuit ( ). These inputs are integrated according to equations (1) and (2):
). These inputs are integrated according to equations (1) and (2):
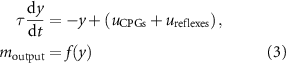
and generate the MN output  . ILPSO, GMAX, and HAMS also receive inputs from the balance controller (
. ILPSO, GMAX, and HAMS also receive inputs from the balance controller ( ). To avoid the activity of the balance controller from being inhibited by the other neural circuits possibly preventing the correct balance of the trunk,
). To avoid the activity of the balance controller from being inhibited by the other neural circuits possibly preventing the correct balance of the trunk,  is not integrated into the MN dynamics, and the final MN output for hip muscles
is not integrated into the MN dynamics, and the final MN output for hip muscles  is defined by the following equation:
is defined by the following equation:
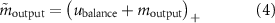
where  is the MN output resulting from the integration of
is the MN output resulting from the integration of  and
and  and
and  represents the balance controller effect on the hip muscles. The operator
represents the balance controller effect on the hip muscles. The operator  represents only the positive part of the selected signal. The amplitude of all components is regulated by the controller's parameters tuned by the optimization algorithm. The muscle activation a responds to the excitation
represents only the positive part of the selected signal. The amplitude of all components is regulated by the controller's parameters tuned by the optimization algorithm. The muscle activation a responds to the excitation  (or
(or  for hip muscles) as defined by Thelen [51].
for hip muscles) as defined by Thelen [51].
Figure 2. Control diagram: the bio-inspired controller is composed of the balance controller of the trunk and the spinal controller divided into CPGs and spinal reflexes. The balance controller aims at keeping the balance of the trunk by stimulating the hip muscles' motoneurons, whereas CPGs and spinal reflexes generate rhythmic behavior stimulating all muscles' motoneurons. Reflexes and CPGs are integrated by motoneurons, whereas balance inputs are summed separately. Created with BioRender.com.
Download figure:
Standard image High-resolution imageThe following sections will describe in detail how each neural input is computed ( ,
,  , and
, and  ).
).
The balance controller is the one proposed by Ong et al [41], and it is the only controller part where a state-machine mechanism is present. A PD control strategy is used to activate the hip muscles balancing the forward lean angle of the trunk. ILPSO, GMAX, and HAMS receive inputs from the balance controller during the stance phase. The excitation given by the balance controller to the hip MNs is described in equation (5).
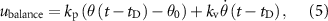
where  and
and  are the proportional and derivative controller's gains, and the constant θ0 is the desired forward lean angle regulating the proportional feedback of the actual forward lean angle θ.
are the proportional and derivative controller's gains, and the constant θ0 is the desired forward lean angle regulating the proportional feedback of the actual forward lean angle θ.  represents the time delay, corresponding to
represents the time delay, corresponding to  ms for the hip muscles. The balance controller has a total of nine parameters.
ms for the hip muscles. The balance controller has a total of nine parameters.
The CPGs were implemented as two coupled oscillators (one per side) composed of RG, and PF layers [36, 46] inspired by the work of Aoi et al [2, 3]. Among the different CPGs models proposed for human locomotion, we chose to take inspiration from Aoi's model [3, 4] to maintain a reasonable level of abstraction for a neural network for which the specific structure is not yet completely clear. Furthermore, the representation of five locomotor synergies is supported by past experimental studies [30].
The RG dictates a periodic command synchronized with the environment through afferents triggered by the heel-strike event. Based on Aoi's model, two coupled differential equations govern the CPG dynamics:
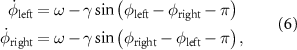
where  denotes the phase of each leg,
denotes the phase of each leg,  is the basic angular frequency, and γ is the coupling constant.
is the basic angular frequency, and γ is the coupling constant.
The differential equation contains events that reset  when the leg touches the ground in order to synchronize the CPGs' phases with the environment. This is the only feedback mechanism present in the CPGs model, and it is described in equation (7):
when the leg touches the ground in order to synchronize the CPGs' phases with the environment. This is the only feedback mechanism present in the CPGs model, and it is described in equation (7):
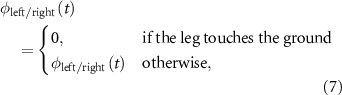
In our simulations, the angular frequency ω has a constant value and represents one of the parameters under optimization.
The pattern formation layer is composed of phase-dependent primitive patterns. Each pattern resembles a bell-shaped waveform with a defined width that can be centered around a specific phase value and is implemented as a raised-cosine function:
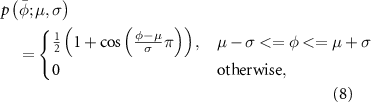
where  is the normalized gait phase, µ is the value corresponding to the peak of the bell shape, and σ is the half-width of the curve. The pattern formation layer is composed of five primitives of the same half-width and centered at different times of the gait phase (figure 3(a)):
is the normalized gait phase, µ is the value corresponding to the peak of the bell shape, and σ is the half-width of the curve. The pattern formation layer is composed of five primitives of the same half-width and centered at different times of the gait phase (figure 3(a)):
Figure 3. CPG structure: (a) the CPG generates five bell-shaped primitives centered at different times of the gait cycle. (b) Each k-pattern stimulates all the m-motoneurons depending on the assigned weight  that can be positive or negative. (a) and (b) Created with BioRender.com.
that can be positive or negative. (a) and (b) Created with BioRender.com.
Download figure:
Standard image High-resolution imageThe choice of modeling the CPG network as the generation of five locomotor primitives has been taken to have smooth and derivable shapes of CPG circuits inputs to MN. More precisely, we took inspiration from the observations done in past experimental studies where five bell-shaped synergies active at different phases of the gait cycle were identified in human studies [30, 31]. However, it should be noted that those recorded synergies account for the overall MN activity and not just the CPG sub-component. Each MN receives a weighted neural excitation or inhibition  from all primitive patterns (figure 3(b)) according to the following equation:
from all primitive patterns (figure 3(b)) according to the following equation:
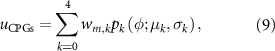
where  is the weight parameter of the pattern k to the MN m to be determined through optimization. The total number of parameters to optimize corresponds to five weights per pattern to each specific muscle and the oscillatory frequency. Therefore, the number of parameters for the CPG network is 48. The possible values assigned to
is the weight parameter of the pattern k to the MN m to be determined through optimization. The total number of parameters to optimize corresponds to five weights per pattern to each specific muscle and the oscillatory frequency. Therefore, the number of parameters for the CPG network is 48. The possible values assigned to  are [−1:1].
are [−1:1].
To implement a physiologically realistic model of sensory-motor control in human locomotion, we model and investigate five spinal reflexes:
Ia afferents provide monosynaptic excitation to MNs innervating the same muscle and disynaptic inhibition mediated by Ia inhibitory INs to antagonistic MNs (figure 4(a)) and model the velocity-dependent response to stretch [11].II afferents provide disynaptic excitation to MNs innervating the same muscle and disynaptic inhibition to antagonistic MNs mediated by excitatory and inhibitory INs, respectively (figure 4(b)). This reflex models the excitatory role of group II afferents [35] responding to changes in muscle length during stretch.Ib afferents provide disynaptic inhibition to MNs innervating the same muscle mediated by inhibitory INs. These INs reciprocally inhibit antagonistic Ib-IN (figure 4(c)). This reflex is triggered by the Golgi tendon organs and it is introduced to protect muscles when large forces are detected [9]. Additionally, Ib afferents provide disynaptic excitation to extensor MNs innervating the same muscle mediated by excitatory INs. These connections model the positive force feedback reversal commonly observed in experimental studies [23, 44, 46] (figure 4(d)).Renshaw cells (RCs) are inhibitory INs providing inhibitions to MNs and Ia-INs innervating the same muscle. Additionally, these cells reciprocally inhibit with antagonistic RCs [55] (figure 4(e)). RCs are activated by MNs innervating the same muscle through synaptic excitation inhibiting these MNs when a large activity is detected to prevent excessive muscle activation.
Comments (0)