
Remember me


Resilience is defined as “the process and outcome of successfully adapting to difficult or challenging life experiences.”1 For those in high stress careers, such as first responders, health workers, surgeons, and military personnel, resilience may be essential. Studies determining risk factors for posttraumatic stress disorder (PTSD), have suggested there are conditioned biological and psychological components that may predisopose individuals to development of PTSD.2–6 While animal studies have explored the biobehavioral expressions of PTSD-resilient and PTSD-susceptible phenotypes in response to trauma,7,8 the present research aims to explore immunological biomarkers for potential detection of resilience, prior to stress exposure. The detection of salivary immune biomarkers that correlate with resilience may help to identify susceptible individuals prone to the development of PTSD and other stress-related breakdowns.
Prior studies have provided insight into mechanisms by which acute stress may modulate individual disease susceptibilities. Neuropsychological processes can alter cellular communication by changing levels of cytokines and hormones, which interact with the immune system through signaling of lymphoid and myeloid cell lines.9,10 Cytokines have the capacity to modify neurotransmission to change behavior, with prior evidence supporting an association between chronic elevations of inflammatory cytokines and the development of neuropsychiatric disorders.11 Our study sought to determine possible cytokines associated with high resilience in first responders.
The Hardiness Resilience Gauge (HRG) is a recently developed and validated psychometric questionnaire, showing significantly increased predictive power over its predecessor, the Dispositional Resilience Scale.12–14 The HRG measures three facets of resilience: Commitment, Control, and Challenge. Together, these make up the trait of Hardiness, which has been positively correlated with adaptive-coping and negatively correlated with avoidance-coping.15–18 We hypothesized that there are correlations between HRG scores and salivary immune biomarker levels/fluctuations at and between timepoints within the acute stress response. These correlations could potentially serve as identifiers for levels of resilience.
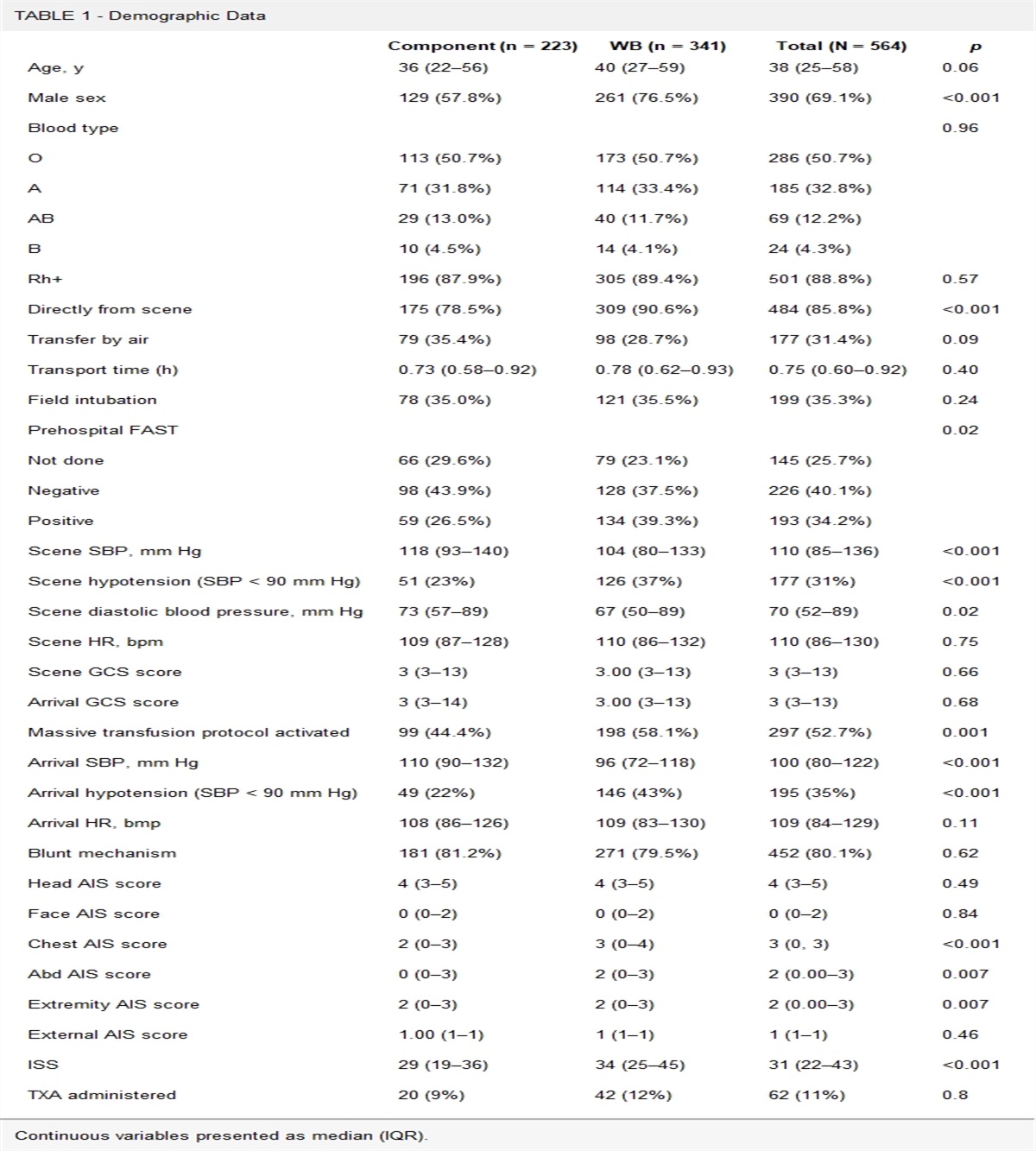
METHODS Study DesignThis is a prospective observational cohort study of 63 consenting first-responders enrolled in a fire academy course in which a cross-sectional design with repeated measures was conducted. Institutional review board approval was obtained (2019-0092). The STROBE statement guidelines were used to ensure proper reporting of methods, results, and discussion (Supplemental Digital Content, SDC 1, https://links.lww.com/TA/D94). Participants provided demographic data and completed the Initial HRG questionnaire during recruitment, at which demographic information was also collected (Table 1). Five days following recruitment, participants underwent a standardized acute stress event, and provided salivary samples at three points in proximity to the event: pre-stress, and recovery. The final HRG was obtained within 12 hours following completion of the stress event. Participants were informed both verbally and in writing about the study content and the measures that would be taken, and each participant gave his or her written informed consent before participating in the study. Participants were excluded for failure to complete the stress event or for missing salivary samples. Outliers were not excluded, on the justification that outliers may be resilience phenotypes of interest.
TABLE 1 - Demographics of Participants Demographics Overall n 59 Age, mean [range] 29.5 [19–48] Sex, n (%) Female 5 (8.5) Male 54 (91.5) Race, n (%) Asian 1 (1.7) Black 3 (5.1) Chinese 1 (1.7) Hispanic 3 (5.1) Indian 1 (1.7) Mixed 2 (3.4) White 48 (81.4) Initial HRG, n (missing) 59 (7) Final HRG, n (missing) 42 (17)Designed to simulate a realistic occupational stressor in a safe environment, the stress test consisted of a physically and cognitively challenging scenario of blindly navigating through a simulated collapsing structure while continuously wearing air masks. This stress test attempts to control the potential influence of participants' demographics and prior stress experiences by using a novel challenge, which is only administered once as a requirement for employment following completion of a fire academy. The study was performed in two separate but comparable cohorts: Academy 1 was conducted in April 2020 and Academy 2 in June 2020. The stress-events for both academies were administered with the same controlled environment and the duration was standardized for each participant. Salivary samples were collected at prestress, poststress, and recovery.
Hardiness-Resiliency GaugeThe HRG was administered through MHS (Mental Health Systems), with the initial questionnaire collected at recruitment, and the final questionnaire collected following completion of sampling. Scores were provided by MHS for the subcategories of control, commitment, and challenge. Details of the specifics of the HRG instrument are published elsewhere.13 Hardiness was determined by the average of these scores for each participant.
Salivary BiomarkersFor saliva sample collection, individuals were given sugar free gum to chew for 5 minutes, after which 1.0 mL of saliva was collected and pipetted into 1.5 mL Eppendorf tubes. Following the collection of salivary samples, a 1 μg/mL concentration of protease inhibitor (20–201 Milipore) was added to the samples. The samples were stored on ice on site until they were shipped on dry ice to determine concentrations using the bead-based HD-42 cytokine plex panel and the HD-6 steroid and thyroid hormone plex assays from Eve Technologies (Alberta, Canada). Cytokines and hormones were reported in units of pg/μL and μg/mL, respectively. Sensitivities of the assays can be found in supplementary material.19
Statistical Analysis/ScoringBecause of the unequal sample sizes and partial sample overlap between the initial and final HRG, a Mann-Whitney U test was employed to nonparametrically compare each subcategory of the initial HRG to the respective final HRG subcategory. Pearson correlation coefficients (r) were calculated with available data separately for either HRG group (initial or final) against biomarker levels/fluctuations within all saliva samples at and between each timepoint, including prestress, poststress, recovery, prestress versus poststress, poststress versus recovery, and prestress versus recovery. To control for false-positives with multiple comparisons, a false discovery rate (FDR) correction was applied to the p values with an α level of 0.05 using the Benjamini-Hochberg procedure.20 All reported results are based upon the FDR-corrected p values. The p values before and after the FDR correction can be found in Supplemental Table 1, https://links.lww.com/TA/D95. Statistical significance was defined by FDR-corrected p values <0.05 and r values ≥0.30 or ≤ −0.30.
RESULTS ParticipantsInitially 63 consenting participants were recruited. Exclusion criteria were met for the following four participants: one did not attend due to COVID-19 diagnosis, two sustained injuries prior to stress-event completion, and one failed to provide saliva samples. Demographics, including age, sex, and ethnicity of the remaining participants, can be found in Table 1 (n = 59). Initial HRG data were missing for seven participants, while Final HRG data was missing for 17 participants.
HRG Score TrendsEvaluation of the spread and mean Initial and Final HRG scores can be seen in Figure 1, which was constructed to analyze the frequency of scores in each subcategory and overall hardiness. Scores for all subcategories were on average higher for the Initial HRG than the Final HRG (Supplemental Table 2, https://links.lww.com/TA/D95). However, the Mann-Whitney U test showed no significant differences between Initial and Final HRG distributions (Supplemental Table 2, https://links.lww.com/TA/D95). The distributions of all subcategories were left-skewed, the means were less than the medians, and the commitment, control, and hardiness subcategories of the final HRG had positive kurtosis, or more data points in the tail than a normal distribution, unlike those of the initial HRG (Fig. 1, Supplemental Table 3, https://links.lww.com/TA/D95).
 Figure 1:
Figure 1: Frequency distributions of Initial and final HRG scores cumulatively (hardiness) and for each subcategory: (A) hardiness, (B) challenge, (C) commitment, (D) control.
Correlation of HRG Scores and CytokinesPearson r correlations were computed for every combination of HRG group (final or initial), HRG subcategory (challenge, commitment, control, or hardiness), and the 48 biomarker levels and fluctuations measured at and between prestress, poststress, recovery. A total of 2,304 correlations were calculated, of which 235 had an uncorrected p value<0.05. Following the FDR correction for multiple comparisons, this was reduced to 59 significant correlations. All significant correlations, with uncorrected and corrected p values, along with sample sizes of each correlation, are shown in Supplemental Table 1, https://links.lww.com/TA/D95.
Initial HRG ResultsFigure 2 displays the correlation heatmap of Initial HRG scores to biomarker levels at prestress, poststress, and timepoints. Figure 3 displays the correlations of Initial HRG scores to biomarker fluctuations between prestress versus poststress, poststress versus recovery, and prestress versus recovery. No significant correlates (p < 0.05; r ≥ 0.30 or ≤ − 0.30) were found between Initial HRG scores and biomarker levels at prestress, poststress, and recovery. Scores on the commitment subcategory of the HRG negatively correlated with change in cortisol levels from poststress versus recovery (Fig. 3B; Pearson's correlation coefficient, r = −0.47; p = 0.029).
 Figure 2:
Figure 2: Correlation heatmaps between biomarker levels and initial HRG scores at prestress (A), poststress (B), and recovery (C) time points (p < 0.05). Significant FDR-corrected correlations (p < 0.05; r ≤ −0.30 or r ≥ 0.30) are indicated with their respective r values.
 Figure 3:
Figure 3: Correlation heatmaps between biomarker fluctuations and initial HRG scores for (A) prestress to poststress, (B) poststress to recovery, (C) prestress to recovery (p < 0.05). Significant FDR-corrected correlations (p < 0.05; r ≤ −0.30 or r ≥ 0.30) are indicated with their respective r values.
Final HRG ResultsCorrelations between Final HRG scores and biomarker levels at prestress, poststress, and recovery are illustrated in Figure 4, and correlations with biomarker fluctuations between prestress versus poststress, poststress versus recovery, and prestress versus recovery are illustrated in Figure 5.
 Figure 4:
Figure 4: Correlation heatmaps between biomarker levels and final HRG scores at prestress (A), poststress (B), and recovery (C) time points (p < 0.05). Significant FDR-corrected correlations (p < 0.05; r ≤ −0.30 or r ≥ 0.30) are indicated with their respective r values.
 Figure 5:
Figure 5: Correlation heatmaps between biomarker fluctuations and final HRG scores for (A) prestress to poststress, (B) poststress to recovery, (C) prestress to recovery (p < 0.05). Significant FDR-corrected correlations (p < 0.05; r ≤ −0.30 or r ≥ 0.30) are indicated with their respective r values.
Scores on commitment negatively correlated with levels of EGF, GROα, interleukin (IL)1Ra, PDGFAA, transforming growth factor α (TGFα), TNFα, and VEGFA at poststress (Fig. 4B; r < −0.45; p < 0.02), and also negatively correlated with change in levels of EGF, GROα, IL15, IL6, and PDGFAA from prestress versus poststress (Fig. 5A; r ≤ −0.46; p ≤ 0.03), as well as for change in cortisol from poststress versus recovery (r = −0.42; p = 0.04). Commitment scores positively correlated with fluctuations in EGF, GROα, IL1Ra, PDGFAA, TGFα, TNFα, and VEGFA from poststress versus recovery (Fig. 5B; r ≥ 0.43; p ≤ 0.03).
Scores on control negatively correlated with FGF2, IL13, IL18, and TNFα at the prestress timepoint (Fig. 4A; r ≤ −0.44; p ≤ 0.045), and negatively correlated with EGF, GROα, IL1Ra, PDGFAA, TGFα, and VEGFA at the poststress timepoint (Fig. 4B; r ≤ −0.49; p ≤ 0.01). From prestress versus poststress, control positively correlated with IL18 (r = 0.47; p = 0.02) and negatively correlated with EGF, GROα, PDGFAA, TGFα, and VEGFA (Fig. 5A; r ≤ −0.43; p ≤ 0.04). From poststress versus recovery, control positively correlated with EGF, GROα, IL1Ra, PDGFAA, and TGFα (Fig. 5B; r ≥ 0.43; p ≤ 0.04). From prestress versus recovery, control positively correlated with FGF2 and IL18 (Fig. 5C; r ≥ 0.47; p ≤ 0.04).
Hardiness negatively correlated with EGF, GROα, PDGFAA, TGFα, and VEGFA at the poststress timepoint (Fig. 4B; r ≤ −0.45; p ≤ 0.03). From prestress versus poststress, hardiness negatively correlated with EGF, GROα, and PDGFAA (Fig. 5A; r ≤ −0.46; p ≤ 0.04). From poststress versus recovery, hardiness positively correlated with EGF, GROα, IL1Ra, PDGFAA, TGFα, TNFα, and VEGFA. From prestress versus recovery, hardiness positively correlated with IL18 (Fig. 5C; r = 0.50; p = 0.04).
DISCUSSIONMany studies suggest a bidirectional feedback loop between the immune system and the neuroendocrine axis. Cytokines influence neuroplasticity, modulating hormones that affect the hypothalamic pituitary axis, including glucocorticoids, which play a role in protection from proinflammatory cytokines.21 In addition, stress exposure can condition alterations of immunologic signaling in mouse cell lines, influencing adaptivity to subsequent stress.22–24 A similar conditioning in response to stress may also occur in humans and could be related to resilience.
Studies have shown IL1B, TNFα, fractalkine (CX3CL1), IL18, and total proinflammatory cytokines to be associated with PTSD and deployed military personnel.25–28 Levels of IL6 and IFNy in Vietnam combat veterans negatively correlated with PTSD severity, yet positively correlated with resilience.29 Patterns of cytokine changes have been demonstrated to vary by time relative to an acute stressor.19 Our study set out to explore specific salivary biomarkers of resilience within these patterns of cytokine changes.
No study, to our knowledge, has attempted to determine the immune response profile to acute stress associated with resilience. A previous study,19 demonstrated a stress-induced salivary inflammatory response in first responders, with noted fluctuations of various cytokines during critical windows of the acute stress response. Our study has found links between cytokine levels during acute stress, and resilience in first responders.
Overall, from initial to final HRG, the HRG scores trended toward a tighter distribution, with mean scores decreasing from Initial HRG to final HRG. This may be due to unequal sample sizes from initial to final HRG, with partial overlap in the samples, which necessitated use of the Mann-Whitney U test. A possible explanation for the decrease in mean scores may be a lack of time to integrate the stressful experience prior to completing the final HRG. Of note, FDR-corrected significant correlations with biomarker levels were primarily found with the final HRG. This may indicate that the HRG becomes more biometrically associated, or that acute salivary biomarker changes become more associated with the HRG, when the HRG is administered more proximal to an acute stress response. There are 13 biomarkers with FDR-corrected significance (Figs. 1-5), of which many correlated with multiple HRG subcategories at specific timepoints. In order of the greatest number of correlations to least, are: EGF, PDGFAA, GROα, TGFα, VEGFA, IL1Ra, TNFα, IL18, cortisol, FGF2, IL13, IL15, and IL6.
Total resilience or hardiness as assessed by the HRG, is inversely related to changes in EGF, GROα, and PDGFAA from prestress to poststress, as well as with levels of those cytokines, TGFα, and VEGFA at poststress. These same cytokines correlate similarly to scores in commitment and control. In addition, increases in these five cytokines, along with IL1Ra and TNFα, from poststress versus recovery were associated with higher resilience. Lastly, increases in the level of IL18 from prestress versus poststress were associated with higher control scores and higher scores in hardiness and control in prestress versus recovery.
Interestingly, EGF, PDGFAA, and GROα followed the same pattern of correlations as each other. These three cytokines had nine significant correlations with the final HRG measurements (Figs. 4 and 5) but no significant correlations with initial HRG measurements (Figs. 2 and 3). Each negatively correlated with commitment, control, and hardiness scores at poststress, suggesting low levels of these cytokines after acute stress are indicative of high resiliency overall and in those respective subcategories (Fig. 4B). While their fluctuations were negatively correlated with commitment, control, and hardiness during acute stress (Fig. 5A; prestress vs. poststress), they were positively correlated during the recovery from acute stress (Fig. 5B; poststress vs. recovery). These results suggest that EGF, PDGFAA, and GROα play a dynamic and potentially interlaced role in resiliency, as increases during acute stress was associated with low resiliency scores while increases during recovery from acute stress was associated with high resiliency.
Other interesting findings included decreases in TGFα and VEGFA from prestress versus poststress associated with higher control scores. Also, in prestress versus recovery, increases in FGF2 were associated with higher control and increases in IL18 were associated with higher control and hardiness. At prestress, FGF2, IL13, IL18, and TNFα are associated with higher control scores. At poststress in addition to the results stated above for EGF, GROα, PDGFAA, TGFα, and VEGFA, low levels of these cytokines in addition to IL1Ra were associated with higher commitment. IL1Ra by itself was also associated with higher control. Interestingly, cortisol was the only biomarker with a significant correlation with the Initial HRG measures, in which decreases in cortisol from poststress versus recovery were associated with increased commitment in both the initial and final HRG measurements.
Epidermal growth factor (EGF), vascular endothelial growth factor (VEGF), and platelet-erived growth factor AA (PDGFAA) all correlated significantly with resilience scores. These are implicated in proliferation of epithelium, vasculature, and mesenchyme, respectively, along with other roles such as brain plasticity and neurogenesis. Circulating levels of EGF and VEGF were found to be significantly lower in patients with exhaustion disorder (ED) compared with healthy controls, suggesting that decreased levels after long-term stress exposure could indicate increased risk of neuronal damage and cognitive impairments. Vascular endothelial growth factor has also been studied for its antidepressant action and ability to operate as a key modulator in the regulation of neurogenesis in the adult hippocampus.30 This study identified that decreases during stress and increases after stress in EGF, GROα, and PDGFAA are protective and predictive for high resilience. In addition, increases in VEGFA following stress, during the recovery process, were also protective and predictive for high resilience.
Groα (CXCL1) was found to follow the same patterns in correlation of EGF and PDGFAA. In a previous study, it was found that elevated levels of Groα at stress were associated with low resilience scores.31 Previous work has shown that major depressive disorder especially in adolescents and elderly patients, is associated with a reduction of CXCL1 in the prefrontal cortex.32 The results of our study suggest GROα is negatively associated with resiliency during stress; however, it is positively associated with resiliency if it is found to increase during the recovery from stress, which suggests a potential dynamic role of this immune marker during acute stress and possible habituation.
One marker found in this study to be associated with lower resilience is TGFα. This protein is produced in macrophages, brain cells, and keratinocytes, inducing epithelial development and stimulating neural cell proliferation in the injured adult brain.33 In animals with reduced expression of TGFα, this causes changes in noradrenaline, serotonin, dopamine, and glutamate, all regulators of the stress response.34 Our study demonstrated that elevated levels and movement early in the stress response was maladaptive, while movement in the recovery from stress was associated with higher resilience. This adds another negative component of sustained stress but suggests a potential protective factor or predictor of higher resilience when levels of TGFα increase in recovery from stress.
A biomarker of note in this study is IL15. A genome-wide study of soldiers diagnosed with PTSD found one region, 4q31, which is close to the gene coding for IL15, associated with PTSD. IL15 is a cytokine involved in the activation and proliferation of T-cell and natural killer cells.27 Our results showed decreases in IL15 in prestress versus poststress conferred higher resiliency in commitment.
Overall, these results suggest a downward trend of immune cytokines from prestress versus poststress, except for IL18, and that lower levels of cytokines in prestress and poststress are predictive for higher resilience scores. Compared with an upward trend of cytokines from poststress versus recovery and prestress versus recovery are predictive for higher resilience scores. These results imply potential advantages if the immune system is less reactive to stress early in the response but more reactive in the 1-hour period of recovery from stress, indicating that a delayed release of cytokines during acute stress may reflect a more conditioned, resilient response. In addition to cortisol's known role in the stress response, this study suggested that sustained release of cortisol in the recovery from stress could be maladaptive for resilience (Fig. 3B & 5B). Cortisol having been the only significant correlate in both initial and final HRG scores may suggest that the decrease in Cortisol following acute stress is an indicator that the HRG test is valuable for measuring resiliency even if taken prior to stress events. Further research should be done with greater sample sizes and varying periods after stress events are needed to establish if other correlates in Final HRG are consistently expressed or if they are dependent on the time at which the HRG measurements are acquired. Exceptionally, there were no immune markers associated with resilience in the challenge subcategory for any of the timepoints or fluctuations. Noteworthy, challenge was the only distribution with negative kurtosis, or less data points in the tails than a normal distribution, in the final HRG. This may be due to a small sample size and future studies should be conducted to illuminate the lack of correlations in challenge.
This study has uncovered possible resilience biomarkers associated with the immune response to stress. The immune system may act as a biological system through which hardiness can have beneficial stress-buffering effects. Our findings have generated a resiliency map (Figs. 2-5) that may help to serve as a screening or diagnostic tool in the future. Future studies should focus on training methods to increase immune markers associated with resilience or pharmacologically modulating levels to attain optimal performance and health during stress.
LIMITATIONSThere are limitations of note in this study. With a relatively small sample size of n = 59, some with missing datapoints, there may not be sufficient power to detect small to moderate differences between groups. In addition, most participants in this study were males and many had prior EMS or military experience, which is not reflective of the general population. This study should serve as a preliminary investigation in resiliency and expanded to a larger group with more diversity in demographics and background. Also, future experiments could examine possible differences in resilience biomarkers in males versus females. Further, the emotional stress of the exercise may have had effects on biomarker levels prior to the stress-event that warrant investigation in futures studies by obtaining samples earlier in the event and/or by recruitment of a non-stress control group. Of note, these analyses were performed with a unique stress test scenario and there may be effects on biomarkers independent of stress (e.g., wearing masks, smoke inhalation, time of day, temperature, activity levels, etc.). Future investigation of these same salivary biomarkers under diverse types of stress is warranted. Salivary samples and select immune panels may not elucidate all the biochemical pathways involved in physiologic and psychological resiliency. The period at which we collected samples may not directly reflect the dynamic changes in biomarker concentrations due to varying durations of activity, transcription, degradation, and release. We recognize that stimulated saliva collection and the lack of standardization in salivary flow rates establish room for potential error in biomarker quantification. The use of HRG scores administered within 1 week of one another also presents a potential confounding effect of memory on scores when comparing initial and final HRG scores. By using Pearson r correlations, we are assuming there are linear relationships between the variables and that the variables are normally distributed and free of outliers. In addition, the FDR correction to reduce false-positives may exclude some true positives.
FUTURE DIRECTIONSFuture investigation of the molecular mechanisms and signaling pathways governing our discovered biomarkers associated with resiliency is justified. This may allow for development of screening protocols for prevention of stress-related breakdowns in susceptible individuals.
AUTHORSHIPC.W. and S.P. contributed to the literature review, data analysis, data interpretation, graphic design of figures and tables, writing, and critical revision. R.R. contributed to the literature review, study design, data collection, supervision, writing, and critical revision. A.L. contributed to conception and study design. K.D.G. contributed to writing and critical revision. F.T. contributed to data interpretation and critical revision. P.B. contributed to data interpretation and critical revision.
ACKNOWLEDGMENTSThis research was supported by an intramural grant through Rocky Vista University. We thank Training Chief Jake Mayhew for allowing us to work with South Metro Fire recruits. We thank Isain Zapata for helping with sample collection.
DISCLOSUREThe authors declare no conflicts of interest.
REFERENCES 1. APA Dictionary of Psychology. https://dictionary.apa.org/. Published February 14, 2022. Accessed September 3, 2022. 2. Yehuda R. Risk and resilience in posttraumatic stress disorder. J Clin Psychiatry. 2004;65(Suppl 1):29–36. 3. Zovkic IB, Sweatt JD. Epigenetic mechanisms in learned fear: implications for PTSD. Neuropsychopharmacology. 2013;38(1):77–93. 4. Meewisse ML, Reitsma JB, De Vries GJ, Gersons BP, Olff M. Cortisol and post-traumatic stress disorder in adults: systematic review and meta-analysis. Br J Psychiatry. 2007;191(5):387–392. 5. Jovanovic T, Kazama A, Bachevalier J, Davis M. Impaired safety signal learning may be a biomarker of PTSD. Neuropharmacology. 2012;62(2):695–704. 6. Eagle AL, Knox D, Roberts MM, Mulo K, Liberzon I, Galloway MP, et al. Single prolonged stress enhances hippocampal glucocorticoid receptor and phosphorylated protein kinase B levels. Neurosci Res. 2013;75(2):130–137. 7. Colucci P, Marchetta E, Mancini GF, Alva P, Chiarotti F, Hasan MT, et al. Predicting susceptibility and resilience in an animal model of post-traumatic stress disorder (PTSD). Transl Psychiatry. 2020;10(1):243. 8. Yehuda R, LeDoux J. Response variation following trauma: a translational neuroscience approach to understanding PTSD. Neuron. 2007;56(1):19–32. 9. Arzt E, Stalla GK. Cytokines: autocrine and paracrine roles in the anterior pituitary. Neuroimmunomodulation. 1996;3(1):28–34. 10. Szałach ŁP, Lisowska KA, Cubała WJ. The influence of antidepressants on the immune system. Arch Immunol Ther Exp. 2019;67(3):143–151. 11. Felger JC, Lotrich FE. Inflammatory cytokines in depression: neurobiological mechanisms and therapeutic implications. Neuroscience. 2013;246:199–229. 12. Bartone PT. Development and Validation of a Short Hardiness Measure. June 1991. http://www.hardiness-resilience.com/docs/aps91b.pdf. 13. Bartone PT, McDonald K, Hansma BJ, Stermac-Stein J, Escobar EMR, Stein SJ, et al. Development and validation of an improved hardiness measure: the Hardiness Resilience Gauge. Eur J Psychol Assess. 2022;19. 14. Windle G, Bennett KM, Noyes J. A methodological review of resilience measurement scales. Health Qual Life Outcomes. 2011;9(1):8. 15. Bartone PT, Homish GG. Influence of hardiness, avoidance coping, and combat exposure on depression in returning war veterans: a moderated-mediation study. J Affect Disord. 2020;265:511–518. 16. Kamtsios S, Bartone P. Preliminary EVALUATION of the psychometric properties of the “hardiness-resilience gauge” in an undergraduate sample. Hell J Psychol. 2021;18(3):287–310. 17. Thomassen ÅG, Hystad SW, Johnsen BH, Johnsen GE, Bartone PT. The effect of hardiness on PTSD symptoms: a prospective mediational approach. Mil Psychol. 2018;30(2):142–151. 18. Vagni M, Maiorano T, Giostra V, Pajardi D, Bartone P. Emergency stress, hardiness, coping strategies and burnout in health care and emergency response workers during the COVID-19 pandemic. Front Psychol. 2022;13:918788. 19. Ryznar R, Wong C, Onat E, Towne F, LaPorta A, Payton M. Principal component analysis of salivary cytokines and hormones in the acute stress response. Front Psychol. 2022;13. 20. Benjamini Y, Yekutieli D. The control of the false discovery rate in multiple testing under dependency. Ann Stat. 2001;29(4):1165–1188. 21. Silverman MN, Pearce BD, Biron CA, Miller AH. Immune modulation of the hypothalamic-pituitary-adrenal (HPA) axis during viral infection. Viral Immunol. 2005;18(1):41–78. 22. Bains JS, Sharkey KA. Stress and immunity—the circuit makes the difference. Nat Immunol. 2022;23(8):1137–1139. 23. Cohen H, Ziv Y, Cardon M, Kaplan Z, Matar MA, Gidron Y, et al. Maladaptation to mental stress mitigated by the adaptive immune system via depletion of naturally occurring regulatory CD4 + CD25+ cells. J Neurobiol. 2006;66(6):552–563. 24. Fleshner M. Stress-evoked sterile inflammation, danger associated molecular patterns (DAMPs), microbial associated molecular patterns (MAMPs) and the inflammasome. Brain Behav Immun. 2013;27(1):1–7. 25. Baumert J, Lukaschek K, Kruse J, Emeny RT, Koenig W, von Känel R, et al. No evidence for an association of posttraumatic stress disorder with circulating levels of CRP and IL-18 in a population-based study. Cytokine. 2013;63(2):201–208. 26. Wang W, Wang L, Xu H, Cao C, Liu P, Luo S, et al. Characteristics of pro- and anti-inflammatory cytokines alteration in PTSD patients exposed to a deadly earthquake. J Affect Disord. 2019;248:52–58. 27. Wang Y, Karstoft K-I, Nievergelt CM, Maihofer AX, Stein MB, Ursano RJ, et al. Post-traumatic stress following military deployment: genetic associations and cross-disorder genetic correlations. J Affect Disord. 2019;252:350–357. 28. Zhang L, Hu XZ, Li X, Chen Z, Benedek DM, Fullerton CS, et al. Potential chemokine biomarkers associated with PTSD onset, risk and resilience as well as stress responses in US military service members. Transl Psychiatry. 2020;10(1):31. 29. Bruenig D, Mehta D, Morris CP, Lawford B, Harvey W, McD Young R, et al. Correlation between interferon γ and interleukin 6 with PTSD and resilience. Psychiatry Res. 2018;260:193–198. 30. Fournier NM, Duman RS. Role of vascular endothelial growth factor in adult hippocampal neurogenesis: implications for the pathophysiology and treatment of depression. Behav Brain Res. 2012;227(2):440–449. 31. Chai H-H, Fu X-C, Ma L, Sun H-T, Chen G-Z, Song M-Y, et al. The chemokine CXCL1 and its receptor CXCR2 contribute to chronic stress-induced depression in mice. FASEB J. 2019;33(8):8853–8864. 32. Korbecki J, Gąssowska-Dobrowolska M, Wójcik J, Szatkowska I, Barczak K, Chlubek M, et al. The importance of CXCL1 in physiology and noncancerous diseases of bone, bone marrow, muscle and the nervous system. Int J Mol Sci. 2022;23(8):4205. 33. Koyama S, Ikeda K, Sato M, Shibahara K, Yuhara K, Fukutomi H, et al. Transforming growth factor-alpha (TGF alpha)-producing gastric carcinoma with acanthosis nigricans: an endocrine effect of TGF alpha in the pathogenesis of cutaneous paraneoplastic syndrome and epithelial hyperplasia of the esophagus. J Gastroenterol. 1997;32(1):71–77. 34. Koshibu K, Levitt P. Transforming growth factor-alpha induces sex-specific neurochemical imbalance in the stress- and memory-associated brain structures. Neuropharmacology. 2006;50(7):807–813.
Comments (0)