
Remember me

Lipid droplets, also known as lipid bodies, are organelles that are ubiquitous and dynamically regulated. They significantly contribute to the production of inflammatory mediators, cell signaling, membrane trafficking, and lipid metabolism [1]. The neutral lipid core of lipid droplets is composed of triacylglycerol, which is synthesized via two major pathways: glycerol phosphate and monoacylglycerol. Sterol esters surround the core, forming a phospholipid monolayer [2]. Three protein families play a major role in triglyceride biosynthesis: glycerol phosphate acyltransferase, acylglycerol phosphate acyltransferase, and Lipin (phosphatidate phosphatase) [3]. Among these, Lipins play a significant role in triglyceride biosynthesis [4]. The Lipin family is evolutionarily conserved across a wide range of organisms, including single-cell eukaryotes, plants, invertebrates, and mammals [5]. Most plants and vertebrates have two Lipin orthologs, while yeast and most invertebrates have only one [6]. In mammals, there are three members of the Lipin family (Lipins 1, 2, and 3), all of which function as magnesium-dependent phosphatidic acid phosphatases. They catalyze the dephosphorylation of phosphatidic acid to diacylglycerol at the endoplasmic reticulum membrane [7]. Lipin plays a crucial role in coordinating the balance between phosphatidic acid and diacylglycerol and also contributes to the formation of triacylglycerol, which is the major form of long-term energy storage in eukaryotes. In addition, Lipin functions as a transcriptional coactivator in a complex with peroxisome proliferator-activated receptor α (PPARα) and peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α) [8]. The complex between Lipin and PGC-1α, PPARα controls the expression of genes that regulate β-oxidation of fatty acids in mitochondria [8]. Previous studies have shown that Lipin 1 promotes protein kinase C activation, which inhibits p21 synthesis and promotes keratinocyte differentiation. Lipins also play a crucial role in cell cycle regulation; knockdown of Lipin 1 arrests the keratinocyte cell cycle in the G1 phase [9]. Furthermore, mutations in Lipin 1 are a common cause of recurrent rhabdomyolysis in children [10].
The Lipin function mentioned above is related to various diseases in humans, such as atherosclerosis, cancer, and obesity [11]. Previous studies have shown that deletion of Lipin can lead to severe chronic somatic muscle tissue lesions in mammals [12]. Homozygous Lipin 1-/-mice exhibit neurological defects, such as tremors, abnormal limb clasping, and abnormal gait [13]. Additionally, Lipins 1 and 2 in the central nervous system (CNS) are associated with TOR1A, which causes dominantly inherited childhood-onset dystonia [13].
Drosophila has a single ortholog of the human Lipin gene, which shows the highest expression in the fat body and ventral ganglion cells in the CNS [7]. Lipins are also expressed in the visceral muscles of the larval midgut and adult testes [7]. Previous studies have shown that Lipin plays essential roles in fat body development and triacylglycerol storage [14]. Knockdown of Lipin perturbs the cell cycle and induces apoptosis during wing development, but the role of Lipin in the nervous system is still under investigation. In this study, a fly model was established by knocking down Lipin in pan-neurons, and the resulting phenotypes were investigated. The obtained data showed that pan-neuron-specific knockdown of Lipin led to reduced locomotor abilities in both the larval and adult stages, accompanied by abnormal morphology at the neuromuscular junction (NMJ). Drosophila is an excellent model for studying human diseases, including neuropathy [15]. This study provides evidence for a direct link between reduced expression of Lipin and neurological phenotypes that could contribute to the pathogenesis of various human diseases in which Lipin is involved.
Materials and methods Drosophila handling and stocksFly stocks were cultured in standard food containing 0.65 wt % agar, 10 wt % glucose, 4 wt % dry yeast, and 5 wt % cornmeal at 25 ˚C. The yellow, white (yw) mutant flies were used as controls. Transgenic flies carrying UAS-Lipin-IR265-272 (77170) and UAS-Lipin-IR277-380 (36007) were obtained from the Bloomington Stock Center (BDSC) at Indiana University and the Vienna Drosophila Resource, respectively. These flies had an inverted repeat of the Lipin gene (targeting regions from amino acids 265 to 272 and 277 to 380, respectively) downstream of the UAS sequence on the third and second chromosomes, respectively. A Gal4 driver strain, elav-Gal4, was also obtained from BDSC.
Quantitative reverse transcription PCRTotal RNA was extracted from 20 third-instar larval CNSs using the standard Qiazol reagent (Qiagen, Hilden, Germany), followed by purification using the Qiagen RNeasy kit. cDNA was synthesized using a SimpliAmpTM Thermal Cycler (Life Technologies, Woodlands, Singapore) according to the manufacturer’s instructions. Quantitative PCR was performed using FastStart Essential DNA Green Master Mix (Roche, Mannheim, Germany) and a LightCycler 96 (Roche). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA was used as an endogenous reference gene. Drosophila whole-genome nucleotide sequences used for primer design were obtained from the FlyBase online database. The following primer sequences were used:
GAPDH F, 5′-GGAGCCACCTATGACGAAATC-3′; GAPDH R: 5′-TCGAACACAGACGAATGGG-3′; Lipin F, 5′-ATCCCACGTCCCTGATATCG-3′; Lipin R, 5′-TTCATCTTGGTTGGTTAGCAGG-3′; Lsd-1 F, 5′- TCTGGAGCAGTTGATTGTGC -3′; Lsd-1 R, 5′- AATATGGTGGGCAACCTCTG -3′; brummer-F, 5′- GGCAATGGGAACAACTGAAC-3′; brummer-R, 5′- TTGATCGGGCAATTTGATGATCC-3′; Hsl-F, 5′- GCAGGAACAACTGATGGAAC-3′; Hsl-R, 5′- GCAACGGGCAATTTGATGATCC-3′.
Production of anti-Lipin antibodyWe purchased the anti-Lipin antibody which was produced by Sigma-Aldrich (Tokyo, Japan) as follows: the Lipin peptide CSTKVDMATLRPIPEK, corresponding to the region from amino acids 1021 to 1035 of the Lipin protein, was conjugated to keyhole limpet hemocyanin (KLH) as a protein carrier. The KLH-conjugated peptide was mixed with Freund’s complete adjuvant and injected into Japanese White rabbits maintained under specific pathogen-free conditions. Antisera were collected from rabbits after 56 days.
Western blottingTotal protein was extracted from ten CNS samples at the third-instar larval stage. The CNS samples were homogenized in sample buffer (150 mM Tris-HCl pH 6.8, 10% SDS, 30% glycerol, 0.01% bromophenol blue, and 0.6 M dithiothreitol). Then, protein extracts were separated on SDS-polyacrylamide gels containing 8% acrylamide and transferred onto polyvinylidene fluoride (PVDF) membranes. After blocking for 1 h with PBST (PBS containing 0.1% Tween-20) and 5% skim milk, the membrane was incubated with rabbit anti-Lipin IgG (1 : 1000) produced in this study for 16 h at 4 ˚C. After washing with PBST, the membrane was incubated with horseradish peroxidase (HRP)-conjugated secondary antibody (1 : 1000, Cell Signaling) in PBST containing 5% skim milk at 25 ˚C for 1 h. Proteins were detected using ECL Select western blotting detection reagents (GE Healthcare, Chicago, USA), and images were taken using AE-9300H Ez-Capture MG (ATTO, Tokyo, Japan), followed by analysis with ImageJ (NIH, Bethesda, USA). The relative level of Lipin was normalized to α-tubulin.
Lifespan analysesParent flies were cultured at 28 ˚C. Newly emerged male adult flies were collected within 24 h and placed in vials at a density of approximately 20 flies per vial. The flies were then maintained on standard food at 28 ˚C. Vials and food were changed every three days, and dead flies were counted daily. The survival rate was determined as the percentage of flies that survived each day until all flies had died.
Crawling assayThe crawling assay was performed as previously described [16], with some modifications. Third-instar male larvae were collected and washed with PBS to remove traces of food. The larvae were then transferred onto 2% agarose gels in Petri dishes. Larval movements were recorded using a digital camera. The recorded videos were converted into AVI format using an MOV-to-AVI converter (Pazera Jacek, Sosnowiec, Poland), and then analyzed for path shape and distance moved using ImageJ software with the wrMTrck plugin.
Climbing assayThe climbing assay was performed as previously described [17]. Adult male flies were collected and transferred to conical tubes (20 flies per tube). The tubes were tapped to collect flies at the bottom, and the length of time adult flies climbed on the glass wall of the tubes for 5 s was recorded using a digital camera. This procedure was repeated five times. Each fly’s climbing performance was evaluated based on the height achieved, with a score of 0 assigned for heights below 2.0 cm, 1 for 2.0–3.9 cm, 2 for 4.0–5.9 cm, 3 for 6.0–7.9 cm, 4 for 8.0–9.9 cm, and 5 for heights above 10.0 cm. Climbing assays were performed every seven days starting from day 3 after eclosion.
Drosophila activity assayMale adult flies, 1 day after eclosion, were analyzed using the Drosophila Activity Monitor (Trikinetics) placed in a 25 ˚C incubator. The flies were monitored for 5 days under a 12 : 12 h light-dark cycle. The activity recorded for each 1-min bin was determined, and the values measured at the endpoint of the 1-min measurement determined the average activity bouts of the flies. Thirty-two adult flies were included in each study group.
ATP level analysisATP levels were assayed using an ATP determination kit (A22066; Molecular Probes; Invitrogen, Thermo Fisher Scientific, Waltham, Massachusetts, USA). Thirty adult flies at 1-day post-eclosion were homogenized in 200 µl of lysis buffer (100 mM Tris, pH 7.8, containing 6 M guanidine HCl and 4 mM disodium EDTA). The samples were boiled for 5 min and centrifuged (10 min, 12 000 × g). The supernatants were then diluted 1 : 100 with lysis buffer, reacted with reagents of the ATP determination kit, and luminescence was measured using a GloMax 20/20 luminometer (Promega, Wisconsin, USA). ATP levels were expressed as luminescence in relative light units.
Triacylglycerol quantificationThirty heads from adult flies, 1 day after eclosion, were homogenized in 100 µl of 0.3% Triton X-100 in PBS. The samples were heated at 90 ˚C for 5 min and chilled on ice for 3 min. After centrifugation at 12,000 × g for 10 min at 4 ˚C, the supernatants were used to determine triacylglycerol contents using a LabAssay Triglyceride kit (Wako Pure Chemical, Osaka, Japan). The absorbance was measured at 600 nm using a microplate reader (SH-1200; Ibaraki, Japan). The triacylglycerol content was calculated using a standard curve made with commercially available triacylglycerol from a standard solution (LabAssay Triglyceride kit, Wako, Tokyo, Japan) and normalized to protein amounts determined using the Pierce BCA assay kit (23225, Thermo Fisher Scientific).
Central nervous system stainingThe CNSs of third-instar larvae were dissected in PBS. Approximately 20 brains were fixed with 4% paraformaldehyde in PBS and stained with antibodies as described. Primary antibodies used included rabbit anti-Lipin antibody produced in this study and rat anti-elav IgG (7E8A10; Developmental Studies Hybridoma Bank, Iowa City, Iowa, USA; DHSB). Samples were then incubated with antirabbit and antirat secondary antibodies (at a ratio of 1 : 800, Molecular Probes, Invitrogen), respectively, for 2 h at 25 ˚C and rewashed with PBS containing 0.3% Triton X-100. Nuclear staining was performed using 4’, 6-diamidino-2-phenylindole. Lipid droplets were stained using Nile Red. The samples were mounted in Vectashield mounting medium (Vector Laboratories, Tokyo, Japan) and inspected using an Olympus FLUOVIEW FV 10i fluorescence microscope, Tokyo, Japan. Fluorescence intensity was analyzed using NIH ImageJ software.
Visualization of neuromuscular junctionsThe morphology of NMJ in muscle 4 at abdominal segment 3 or 4 in third-instar larvae was used to visualize NMJ, as previously described [18]. The wandering third-instar male larvae were dissected in a modified minimal hemolymph-like 3 saline and fixed with 4% paraformaldehyde in PBS for 40 min at 25 ˚C. The samples were washed with PBS containing 0.3% Triton X-100 and blocked with 10% goat serum in PBS-0.15% Triton X-100 for 30 min at 25 ˚C. Then, the processed samples were incubated with mouse anti-bruchpilot IgG (1 : 100) (nc82, DHSB) for 16 h at 4 ˚C. After being washed with PBS containing 0.3% Triton X-100, the samples were incubated with a goat antimouse secondary antibody labeled with Alexa 594, Cell Signaling, Massachusetts, USA, together with FITC-conjugated goat anti-HRP IgG (1 : 400) (Jackson ImmunoResearch, Pennsylvania, USA) for 2 h at 25 ˚C and then rewashed with PBS containing 0.3% Triton X-100. The samples were mounted using VECTASHIELD (Vector Laboratories). Confocal microscopy images were obtained using a fluorescence FV10i microscope (Olympus FLUOVIEW FV10i) and analyzed the total branch length, the size of the largest bouton, and the number of boutons on the branch using ImageJ software. Because morphology of NMJ in segments 3 and 4 were similar, the average of these obtained values in both segments 3 and 4 was calculated for individual larva.
Statistical analysisAll data were collected by using Microsoft Excel (Microsoft, Washington, USA), statistically analyzed, and graphed by using GraphPad Prism 9.5.1 (GraphPad Software, Boston, USA). The statistical test for each experiment is detailed in the figure legend.
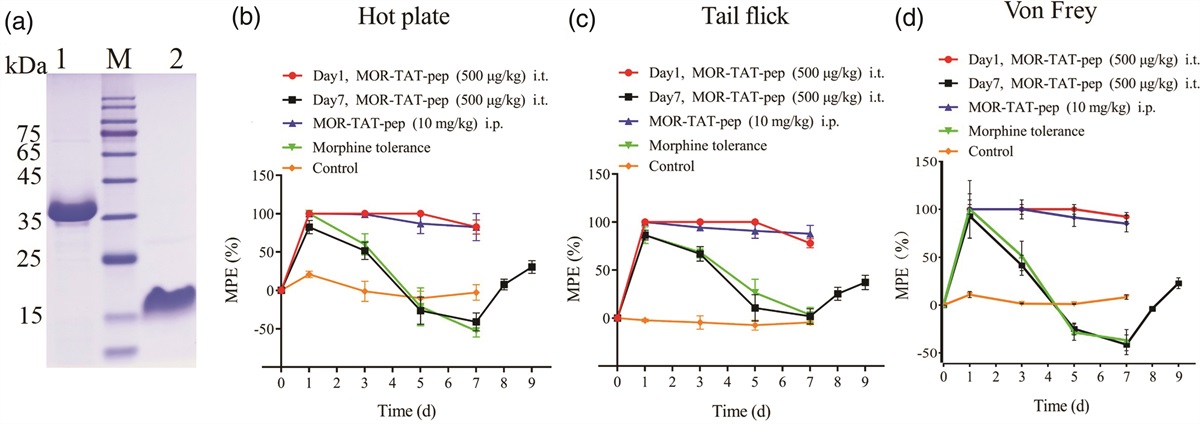
Results Knockdown of Lipin in the larval pan-neuron induces defects in locomotor ability and activity, life span, and circadian activityTo analyze the localization of Lipin in the CNS, we generated an antibody against Lipin and confirmed its specificity by western blot analysis of larval CNS extracts. In control flies (elav > yw), the antibody showed a major band with an apparent molecular mass of 174 kDa (Fig. 1b, lane 1). Using anti-Lipin IgG and anti-Elav IgG, we performed co-immunostaining of CNSs dissected from third-instar larvae. As shown in Fig. 1d, Lipin in yw was detected mainly in the cytoplasm of neurons throughout the central brain and the ventral nerve cord.
 Fig. 1:
Fig. 1: Expression of Lipin protein in the neurons of the larval CNS. (a) The relative expression level of Lipin mRNA in the third-instar larval CNS was determined using RT-qPCR. (b and c) The larval CNS extracts were subjected to western blotting in (b), and the Lipin protein signal intensities were analyzed using ImageJ software in (c). The expression levels of Lipin protein were significantly decreased in both knockdown fly lines. (d) The larval CNS of yw was costained with (a, d, g) anti-Lipin IgG and (b, e, h) anti-Elav IgG, followed by Alexa 488-labeled antirabbit and Alexa 594-labeled antirat, respectively. Merged images are shown in (c, f, i). (a–c) Whole CNS, scale bar: 100 µm; (d–f) Brain lobe and (g–i) ventral nerve cord (VNC), scale bar: 20 µm. (a and c): one-way ANOVA with Dunnett’s multiple comparisons post-hoc test was performed. Data are presented by mean ± SD; ***, P < 0.001, n = 3 biological replicates. Genotypes: elav > yw (yw/+; +; elav-Gal4/+), elav > Lipin-IR265-272 (w/+; UAS-Lipin-IR265-272/+; elav-Gal4/+), and elav > Lipin-IR277-380 (w/+; +; UAS-Lipin-IR277-380/elav-Gal4). CNS, central nervous system; RT-qPCR, quantitative reverse transcription PCR.
To reveal the role of Lipin in neuronal function, we specifically knocked down Lipin in pan-neurons using embryonic lethal abnormal vision (elav)-GAL4, a pan-neuronal GAL4 driver. We confirmed, through quantitative reverse transcription PCR (RT-qPCR) analyses that the mRNA expression of Lipin in two independent knockdown fly lines was significantly decreased to 12.6% (elav > Lipin-IR265-272) and 20.9% (elav > Lipin-IR277-380), respectively, in the CNSs of third-instar larvae (Fig. 1a). The intensities of the major bands in the western blotting of Lipin knockdown flies were decreased to 28.98% (elav > Lipin-IR265-272) and 38.35% (elav > Lipin-IR277-380), compared to that of control flies (Fig. 1b and c). The immunostaining signal also decreased in the CNS of Lipin knockdown third-instar larvae (Supplementary Fig. S1, Supplemental digital content 1, https://links.lww.com/WNR/A703). These data revealed that Lipin was effectively knocked down in the CNS of Lipin knockdown larvae.
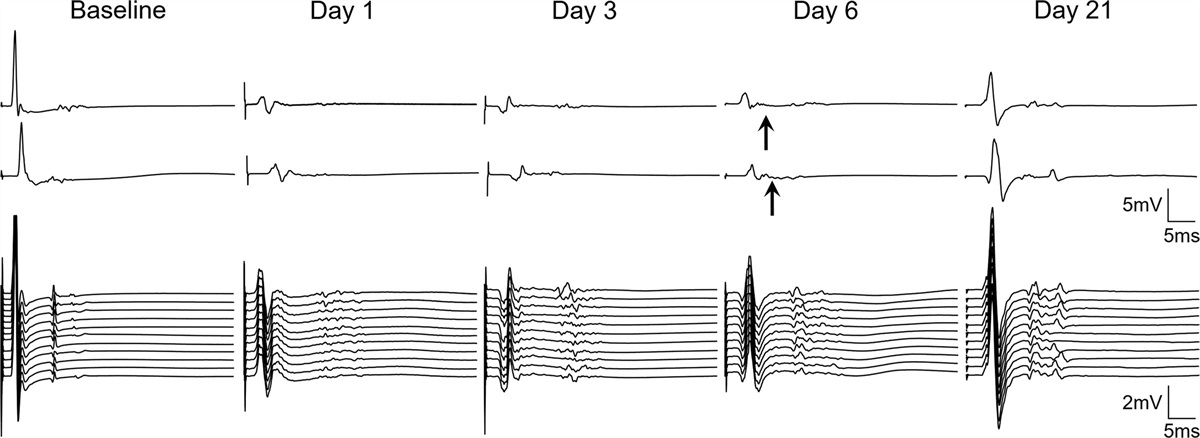
Next, we investigated the effects of pan-neuron-specific knockdown of Lipin on the locomotor behavior of larvae and adults by performing crawling and climbing assays, respectively. Lipin knockdown larvae exhibited significant decreases in crawling distance and speed compared to control larvae (Fig. 2a and b). As shown in Fig. 2b, the average speed of knockdown larvae was significantly reduced in Lipin knockdown flies (elav > Lipin-IR265-272 and elav > Lipin-IR277-380) by 0.31 mm/s and 0.32 mm/s, respectively, in comparison with that of control flies (elav > yw).
 Fig. 2:
Fig. 2: Phenotypes of pan-neuron-specific Lipin knockdown larvae and adult flies. (a and b) Knockdown of Lipin in pan-neurons decreased the crawling ability of larvae. (a) Representative depiction of the larval path. (b) Quantitative data of the average crawling speed. One-way ANOVA with Dunnett’s multiple comparisons post-hoc test was performed. (c) The percentage of surviving flies (elav > yw, elav > Lipin-IR265-272, and elav > Lipin-IR277-380) is shown. The black line indicates the median lifespan. Lipin knockdown flies showed a statistically significant decrease in survival (n = 120). (d) Climbing assay. Lipin knockdown caused a decrease in climbing ability in adults (n = 80 for each genotype). (e–g) Drosophila activity assay. Knockdown of Lipin decreased the activity of flies in the daytime, night time, and all day. The fly genotypes were elav > yw (blue, n = 32), elav > Lipin-IR265-272 (red, n = 32), and elav > Lipin-IR277-380 (green, n = 32. (e) Black or white bars above the graphs indicate light conditions. (b and d) and (f–g): Data are presented by beeswarm and boxplots. (c and e): Time series data are shown by line with error bands with 95% confidence interval. (d) and (e–g): Kruskal–Wallis test with Dunn’s multiple comparisons test; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001). Genotypes: elav > yw (yw/+; +; elav-Gal4/+), elav > Lipin-IR265-272 (w/+; UAS-Lipin-IR265-272/+; elav-Gal4/+), and elav > Lipin-IR277-380 (w/+; +; UAS- Lipin-IR277-380/elav-Gal4).
Although Lipin knockdown larvae exhibited decreased crawling ability, they developed into adult flies normally; however, the median survival time of Lipin knockdown flies (elav > Lipin-IR265-272, 40 days; elav > Lipin-IR277-380, 40 days) was slightly but significantly shorter than that of control flies (43 days) (Log-rank test, P = 0.003) (Fig. 2c). The climbing assay demonstrated that the locomotor abilities of Lipin knockdown adult flies significantly decreased throughout the adult stage (Fig. 2d). Therefore, pan-neuron-specific Lipin knockdown induces deficient locomotor ability at both the larval and adult stages.
It has been reported that lipid storage is closely related to the circadian rhythm [19]. Therefore, we investigated the effect of Lipin knockdown on circadian activity in adult flies. The results are shown in Fig. 2e–g. The activity of daytime was reduced in both elav > Lipin-IR265-272 and elav > Lipin-IR277-380 flies, while activity during the night did not change.
Lipin knockdown reduces ATP levelATP is the primary source of energy for cells, including neurons, and is involved in many cellular processes, such as protein synthesis, ion transport, and nerve impulse propagation [20]. Dysfunctions in the production or utilization of ATP have been linked to various neurodegenerative disorders, including Alzheimer’s and Parkinson’s diseases [21]. Therefore, we hypothesized that Lipin depletion leads to decreased ATP production in the nervous system. To test this hypothesis, we examined ATP levels in both control and pan-neuron-specific Lipin knockdown flies. The ATP levels in the heads of adult flies, elav > Lipin-IR265-272 and elav > Lipin-IR277-380, were decreased by 60.7% and 56.7%, respectively, compared to the level in control flies, elav > yw (Fig. 3e). These findings suggest that Drosophila Lipin plays a role in energy production in the CNS.
 Fig. 3:
Fig. 3: Lipin knockdown reduces lipid contents and ATP levels in Drosophila. (a) Third-instar larval CNSs were stained with Nile red. Scale bar indicates 50 µm. Lipid droplet size and the number of lipid droplets are shown in (b and c), respectively. (d) Triacylglycerol contents of head extracts of 1-day-old adult flies were expressed as the ratio of triacylglycerol/protein (w/w). (e) Quantitative data of ATP levels in head extracts of adult flies are shown. The data are presented as mean ± SD and t-test was performed. *, P < 0.05; **, P < 0.01; ***, P < 0.001. Genotypes: elav > yw (yw/+; +; elav-Gal4/+) and elav > Lipin-IR265-272 (w/+; UAS-Lipin-IR265-272/+; elav-Gal4/+). CNS, central nervous system.
Lipin is involved in triacylglycerol synthesis in the central nervous systemTo examine whether lipid storage is reduced in Lipin knockdown flies, we investigated the size and number of lipid droplets in the larval CNS and the triacylglycerol content in the heads of adult flies. To visualize lipid droplets, the CNS of third-instar larvae was stained with Nile Red. The data indicated that Lipin knockdown reduced the size and number of lipid droplets in the third-instar larval CNS (Fig. 3a–c). In addition, the triacylglycerol level in the heads of adult flies was also decreased to 72.3% in the Lipin knockdown flies compared to that in control flies (Fig. 3d). Collectively, these data indicate that Lipin is required for lipid storage in Drosophila.
Neuron-specific Lipin knockdown affects the morphology and the number of active zones at the neuromuscular junctionBecause neuron-specific Lipin knockdown leads to locomotor defects, we investigated the morphology and function of the NMJ in Lipin knockdown larvae. Previous studies have demonstrated that Drosophila larval NMJ is a well established system for studying synaptic function and morphology [22]. In this study, the total branch length from the intersegmental nerve, the size of the largest bouton, and the number of boutons on the branch in third-instar larvae were measured after visualization using an anti-HRP antibody, a neural membrane marker. The branch length of both knockdown larvae, elav > Lipin-IR265-272 and elav > Lipin-IR277-380, was significantly shorter than that of control larvae (Fig. 4a and c). Moreover, the number of boutons and bouton size was considerably reduced compared to those in the control flies (Fig. 4a, d, e). Boutons contain multiple active zones, where neurotransmitters are released, each of which is opposite to a glutamate receptor cluster. The active zones of the boutons were evaluated using the cell adhesion molecule Bruchpilot, which is the central compartment of the electron-dense cytomatrix in the active zone. As shown in Fig. 4b, f the number of active zones was significantly reduced in Lipin knockdown larvae. These results suggest that Lipin plays an essential role in regulating active zone structure formation at NMJs and that neuronal dysfunction might be caused by decreased synaptic vesicles containing neurotransmitters.
 Fig. 4:
Fig. 4: Knockdown of Lipin causes abnormal morphology of synapse at NMJ of third instar larvae. (a) NMJs were labeled with FITC-labeled anti-HRP IgG, which reacts with an insect neural membrane molecule. NMJs of flies elav > yw (yw/+; +; elav-Gal4/+) (a, d), elav > Lipin-IR265-272 (w/+; UAS-Lipin-IR265-272/+; elav-Gal4/+) (b, e), and elav > Lipin-IR277-380 (w/+; +; UAS-Lipin-IR277-380/elav-Gal4) (c–f) are shown, scale bar: 10 µm (a–c); 5 µm (d–f). (b) NMJs were costained with anti-HRP (a, e, i) and anti-bruchpilot (b, f, j) IgGs. Merged pictures (c, g, k), and region of interest (ROI) (d, h, l) in merged pictures are also shown. NMJ of flies carrying elav > yw (yw/+; +; elav-Gal4/+) (a–d), elav > Lipin-IR265-272 (w/+; UAS-Lipin-IR265-272/+; elav-Gal4/+) (e–h), and elav > Lipin-IR277-380 (w/+; +; UAS-Lipin-IR277-380/elav-Gal4) (i-l) are shown, scale bar: 10 µm. (c) Longest branch length was measured from one arrowhead to another arrowhead in pictures a, b, c. (d) Bouton number. (e) Bouton size. (f) Bruchpilot positive signal number. (c–f): One-way ANOVA with Dunnett’s multiple comparisons post-hoc test was performed. The data are presented as mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001. HRP, horseradish peroxidase; NMJ, neuromuscular junction.
DiscussionPrevious studies have revealed that Lipin is expressed not only in the fat body but also in almost all other tissues, including the testis, gut, intestinal tract/Malpighian tubules, carcass, and CNS [7,14]. Drosophila expresses at least three isoforms of Lipin in specific tissues [7]. In Drosophila, Lipins play an essential role in forming lipid droplets. In the present study, we also detected Lipin in the larval CNS using a specialized antibody, anti-Lipin (Fig. 1), but not in the NMJ (data not shown). This suggests that Lipin mainly plays a role in the CNS and not in the peripheral nervous system in larvae. A previous study in mice showed that mammalian Lipin 1 can localize to either the nucleus or the cytoplasm [8]; however, the present study showed that Drosophila Lipin localizes strongly in the cytoplasm of neurons. This discrepancy suggests that Drosophila Lipin plays a major role in the cytoplasm of neural cells as an enzyme, phosphatidic acid, rather than as a transcriptional coactivator.
To clarify whether the reduction in Lipin expression is critical for defects in neuronal function and the development of neurodegeneration, we performed neuron-specific knockdown of Lipin using elav-Gal4 and analyzed the phenotypes of locomotor ability, longevity, and circadian activity (Fig. 2). We also analyzed the anatomical defects in the presynaptic terminals of motor neurons in third-instar larvae (Fig. 4).
Evidence from human and animal studies has shown that a high-fat diet can affect both circadian rhythms and energy metabolism [23]. In addition, a previous study has reported that knocking down the circadian gene Clock in central clock cells leads to enhanced triacylglycerol accumulation in the fat body. Therefore, triacylglycerols appear to be controlled by central clock neurons [19]. To further understand the relationship between circadian rhythm and lipid formation, the circadian rhythm was checked in both control and Lipin knockdown flies. The circadian rhythm of Lipin-knockdown flies oscillated at an out-of-phase level in the morning, and slight differences were observed in the rhythm. Both Lipin knockdown strains used in this study showed low-amplitude rhythms. These findings with fly models support the hypothesis that circadian rhythms are related to lipid formation and metabolism.
Moreover, the present results revealed a critical role of Lipin in the morphogenesis and function of peripheral neurons at the NMJ, such as a decrease in the total length of the synaptic branches of motor neurons. Drosophila NMJ synapses are glutamatergic, similar to those in the vertebrate CNS. The potential and paramount role of redox signaling is a key aspect of neurodegenerative diseases. Here, we also found that the knockdown of Lipin decreased ATP levels, causing aberrant neuronal morphology and a reduction in the active zone, an indicator of neuronal dysfunction in NMJs, resulting in defects in locomotive ability.
Lipid droplets affect cellular physiology and function in the CNS, with the brain being the second most lipid-rich organ. On the other hand, Lipin is an enzyme responsible for catalyzing the production of diacylglycerol from phosphatidic acid and maintaining a balance between phosphatidic acid and diacylglycerol, which is a crucial substrate for the synthesis of triacylglycerol [10]. Previous studies have demonstrated that mutation of Lipin induces decreased triacylglycerol levels and leads to changes in the organelle structure of the fat body, such as in the cell nucleus, mitochondria, and autophagosome [24]. Although triacylglycerol levels are lower in the CNS than those in the fat body [7], their roles in the integration of organismal triacylglycerol homeostasis and pathophysiological phenotypes in response to triacylglycerol dysregulation remain equally significant [25]. In the present study, the levels of triacylglycerol were significantly decreased in Lipin knockdown flies (Fig. 3d). Furthermore, we checked the expression of two lipases, Bummer and hormone-sensitive lipase (Hsl), using RT-qPCR. The mRNA level of
Comments (0)