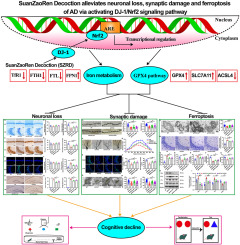
Diabetes-associated cognitive dysfunction (DACD) is a common central nervous system complication of diabetes mellitus (DM), which is clinically manifested by memory deterioration, decreased learning ability, and language, comprehension, and judgment disorders (Biessels and Despa, 2018). In China, 60–70% of DM patients show varying degrees of cognitive dysfunction (Song et al., 2017). It has been found (Klimova et al., 2018; Xue et al., 2019; You et al., 2021) that diabetes has been a major risk factor for increasing the risk of cognitive disorder by 1.25–1.91 times, with severe cases even progressing to irreversible dementia. The autopsy revealed that degenerative changes such as impaired acetylcholine synthesis, β-amyloid (Aβ) deposition, hyperphosphorylation of Tau proteins, and loss of neurons appeared in brain regions with dense distribution of insulin receptors in diabetic patients, such as the hippocampus and part of the cerebral cortex (Sonnen et al., 2009). It has been found (Zhang et al., 2019) that the hippocampus volume of diabetic patients is reduced compared with that of normal people of the same age, and the ultrastructure of neuronal cells in the hippocampal region is disrupted, with apoptotic manifestations, such as nuclear consolidation and cell volume reduction, occurring in advance. Therefore, reducing excessive apoptosis of hippocampal neurons is a key link in the prevention and treatment of DACD.
Autophagy is a highly conserved mechanism of catabolism developed during the evolution of eukaryotes, which plays a regulatory role in intracellular monitoring, scavenging, and supply (Dikic and Elazar, 2018). Studies have shown (Menzies et al., 2017) that non-selective autophagy randomly engulfs intracellular metabolic wastes and senescent components to complete the material and energy cycle, while selective autophagy can degrade dysfunctional organelles, abnormally accumulated aggregates, and specific metabolites to adapt neuronal cells to lipid imbalances, glucose deficiencies, and amino acid deficiencies in response to environmental changes and pathological stimuli, affecting the course of diabetic neurodegenerative diseases. Autophagy levels are closely linked to the deposition of phosphorylated Tau protein, which can promote microtubule assembly and stabilization, playing an essential role in the process of retrograde transport and maturation of autophagosomes and the fusion with lysosomes. Decreased autophagy levels lead to the formation of Tau protein low-molecular polymer and insoluble aggregates, inducing hyperphosphorylation of Tau protein, which then causes lysosomal aberrations and exacerbates autophagy dysfunction, leading to synaptic damage and memory decline (Qu et al., 2011; Caballero et al., 2018). Studies have shown (Jing et al., 2017; Miao et al., 2019) that the compensatory increase in autophagy levels in the early stage of diabetes can play a neuroprotective role. However, with the prolongation of the disease, the decreased levels of cellular autophagy and impeded autophagic flux cause excessive apoptosis of hippocampal neurons, resulting in cognitive dysfunction. At this time, the inhibition of autophagy by administration of 3-methyladenine (3 MA) can exacerbate high glucose-induced neuronal apoptosis, whereas restoration of autophagic flux can antagonize cognitive dysfunction and alleviate the damage to hippocampus organization (Zhou, 2017).
The AMPK/mTOR pathway-mediated autophagy is of great significance to the prevention and treatment of DACD. Activated mTORC1 can inhibit the generation of autophagic precursors, extension of autophagic precursors and the autolysosome formation process (Xu et al., 2022). mTORC1 can phosphorylate the S395 site of WIPI2 to promote the ubiquitylation of WIPI2 and its degradation via the proteasome, affecting the recruitment of ATG12-ATG5-ATG16L and the lipidation modification of LC3 family proteins. It can be used to determine the autophagy intensity. In addition, mTORC1 can phosphorylate the S498 site of UVRAG to inhibit the lipokinase activity of VPS34 complex 2 and the maturation of autophagosomes. Meanwhile, it can inhibit the recruitment of the HOPS complex by UVRAG and autophagosome-lysosome fusion (Chang et al., 2009). AMPK is an energy receptor widely expressed in the brain to activate autophagy, which can directly phosphorylate the key component protein Raptor of the mTORC1 complex and inhibit the small GTP enzyme Rheb necessary for mTORC1 activation by phosphorylating the TSC1/2 complex in tuberous sclerosis, inhibiting the mTORC1 (Egan et al., 2011; Liu, 2019). AMPK signaling is involved in multiple aspects of brain development and function, including neuronal proliferation and migration, morphogenesis, and synaptic growth (Dasgupta and Milbrandt, 2009; Zang et al., 2009; Amato and Man, 2011). It has been found (Kong et al., 2018; Chen et al., 2019; Gu et al., 2019) that metformin, liraglutide, and curcumin nicotinate can intervene in autophagy, attenuate Aβ deposition, and tau phosphorylation, and improve cognitive dysfunction by regulating AMPK molecules and their downstream pathways.
In terms of treating DACD patients, traditional Chinese medicine has the advantages of multicomponent, multitarget, and multi-pathway properties (Meng et al., 2021). Tangzhiqing decoction (TZQD) is a traditional herbal formula composed of five widely applied TCM prescriptions, Huangjing (Polygonatum sibiricum F. Delaroche), Gouqizi (Lycium barbarum L.), Zelan (Lycopus lucidus Turcz. ex Benth.), Guijianyu (Euonymus alatus (Thunb.) Siebold) and Jiangcan (Bombyx batryticatus), developed by Jiangsu Province Hospital of Chinese Medicine for the treatment of diabetes and its complications. TZQD has the effects of tonifying Qi, nourishing Yin, promoting blood circulation, and resolving phlegm. Pharmacological studies have shown that Huangjing (Liu et al., 2021), Guijianyu (Fan et al., 2020), and Gouqizi (Tian et al., 2019) have the effects of regulating blood sugar levels and anti-dementia; Zelan (Zhang et al., 2021) is beneficial for reducing blood lipids and reducing blood clots; Jiangcan (Hu et al., 2017) functions as a blood sugar regulator and neuroprotectant. A series of clinical randomized controlled studies (Zhang et al., 2011; Huang, 2012; Xie, 2013) found that TZQD can effectively reduce blood sugar, blood lipids, BMI, fasting insulin level, and whole blood viscosity, and improve insulin resistance in patients with type 2 diabetes. TZQD plays a potential cognitive protective role in diabetes patients by controlling the risk factors of DACD such as hyperglycemia (Crane et al., 2013), insulin resistance (Arnold et al., 2018), vascular injury (Wardlaw et al., 2019), and obesity (Albanese et al., 2017). It has been found by our research group (Wang et al., 2014, 2019; Yang et al., 2018; Shi, 2022) that TZQD can significantly improve the learning and memory ability of type 2 diabetic mice as well as improve endoplasmic reticulum stress and insulin resistance, and regulate the expression levels of apoptosis-related proteins, Bax, Bcl-2, Caspase 3, and Caspase 8 in hippocampus organization. Meanwhile, the active ingredients of TZQD, such as quercetin (Han et al., 2021), wolfberry polysaccharide (Liu et al., 2018; Dai et al., 2022), and baicalein (Kuang et al., 2017), were found to have strong blood-brain-barrier penetration and regulate autophagy to exert neuron protection in the previous network pharmacological studies (Qiu and Wang, 2021; Shi, 2022). What's more, TZQD prescription can affect the expression of autophagy-related proteins with its protective effect against high glucose-induced HT22 cell damage in vitro, but its specific mechanism remains unclear (Shi et al., 2022).
The study aims to further elucidate the protection of TZQD on cognitive function in mice with T2DM and determine whether the underlying mechanism is related to the regulation of the AMPK/mTOR pathway to activate autophagy.


Comments (0)